熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
Metabolism in anoxic permeable sediments is dominated by eukaryotic dark fermentation
以真核暗發酵為主的缺氧滲透沉積物的代謝
來源:PUBLISHED ONLINE: 28 NOVEMBER 2016
論文摘要
本論文摘要指出,透水性(沙質)沉積物在陸架區廣泛分布,是海洋生物地球化學循環的關鍵貢獻者。這些沉積物中的有機質主要由微藻構成。與細菌和古菌不同,真核生物擁有獨特的厭氧代謝途徑。本研究通過流通式反應器實驗證明,在透水性沉積物的厭氧環境中,溶解無機碳(DIC)的生產主要源于真核生物的厭氧代謝活動。實驗中發現,DIC的厭氧生產始終伴隨著大量的溶解氫氣(H?)產生,表明存在發酵過程。使用廣譜殺菌抗生素后,DIC和H?的生產依然持續,但被甲硝唑(一種特異性抑制真核生物鐵氧還蛋白/氫化酶途徑的藥物)處理后則停止。代謝組學分析顯示,厭氧開始時脂質產量大幅增加。細胞計數顯示沉積物中以微藻為主。從研究點分離的硅藻和綠藻在黑暗厭氧培養中均產生了H?,證實了微藻進行發酵的假說。研究得出結論,微藻的暗發酵可能是透水性沉積物中一種重要的能量守恒途徑,挑戰了傳統上認為細菌過程主導沉積物碳礦化的觀點。
研究目的
本研究的核心目的在于:
識別主導代謝途徑:探究在透水性沉積物的厭氧條件下,是哪種微生物(細菌還是真核微藻)以及哪種代謝途徑(如硝酸鹽還原、硫酸鹽還原還是發酵)主導有機碳的礦化(以DIC產量衡量)。
驗證一個顛覆性假說:挑戰“沉積物厭氧代謝主要由細菌的呼吸鏈(硝酸鹽→鐵→硫酸鹽還原)主導”的傳統范式,驗證 “真核微藻的暗發酵”是否在特定環境中扮演主要角色。
闡明能量守恒機制:理解在缺乏光和氧氣的動態沙質沉積物中,占生物量主導地位的微藻如何生存并獲取能量。研究旨在證明暗發酵是其關鍵的生存策略。
確認H?產生的來源和機制:探究觀測到的高濃度H?的來源,并通過藥理學抑制實驗和純培養驗證,證明其源于真核微藻的特定發酵途徑(鐵氧還蛋白/氫化酶途徑),而非細菌。
研究思路
本研究采用了“宏觀現象觀測 → 機制排除與推定 → 特異性驗證 → 多證據鏈整合”的嚴謹思路:
現場觀測與初始實驗:從澳大利亞菲利普港灣和丹麥凱特明德兩個地點的透水沉積物取樣。使用流通式反應器(FTR)模擬原位條件,監測在厭氧條件下DIC、H?、氮氣(N?)等溶質的產生速率。
排除傳統機制:通過添加硝酸鹽(NO??)作為電子受體,發現其還原速率遠低于DIC生產速率,排除了硝酸鹽呼吸作為主要途徑。同時,鐵還原、硫酸鹽還原和產甲烷的速率也極低,無法解釋觀測到的大量DIC產生。
識別關鍵線索 - H? 的產生:發現厭氧條件下持續產生高濃度H?,其產量遠高于已知的細菌發酵產H?水平,提示存在一個更重要的H?源。
區分生物域貢獻:使用廣譜抗生素(阿莫西林)抑制細菌活性。發現抗生素處理并未抑制DIC和H?的產生,但抑制了典型的細菌過程(反硝化)。這強有力地表明真核生物是主要貢獻者。
確定具體代謝途徑:使用甲硝唑,一種特異性抑制真核生物[FeFe]-氫化酶(暗發酵關鍵酶)的藥物。發現甲硝唑完全抑制了H?的產生,從而將途徑鎖定為真核生物的暗發酵。
代謝物與純培養驗證:
代謝組學:分析顯示厭氧條件下脂質(如油酸酯)顯著增加,這與微藻暗發酵將底物轉化為儲能脂質的已知途徑一致。
純培養實驗:從沉積物中分離出多種硅藻和綠藻,在黑暗厭氧條件下培養,均成功觀測到H?的產生,為假說提供了最直接的證據。
構建概念模型:基于所有證據,提出了一個概念模型(圖3),描繪了微藻在動態沉積物中通過暗發酵維持生存的能量策略。
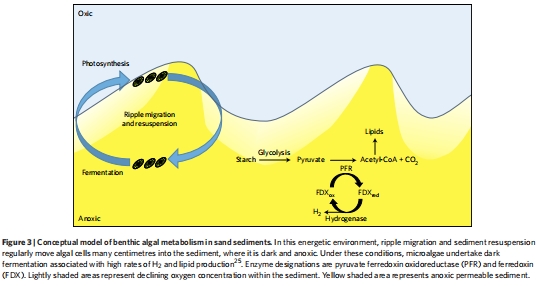
測量數據及其研究意義
研究測量了多方面的數據,其意義和來源如下(數據均引用自文檔中的圖表):
溶質通量速率(量化沉積物代謝強度)
測量指標:通過FTR進口和出口的濃度差計算的溶解無機碳(DIC)產生速率、氫氣(H?)產生速率、硝酸鹽(NO??)消耗速率、氮氣(N?)產生速率等。
研究意義:圖1a, b顯示,在厭氧條件下,DIC的產生速率很高,而硝酸鹽還原等傳統呼吸途徑的貢獻很小(<10%)。圖2a顯示厭氧啟動后約36小時開始出現大量的H?生產。這些數據是整個研究的起點和核心證據,揭示了存在一種不依賴傳統電子受體的、高產H?的強大厭氧代謝。
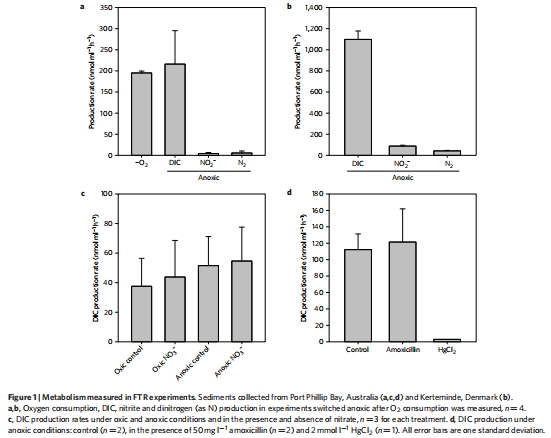
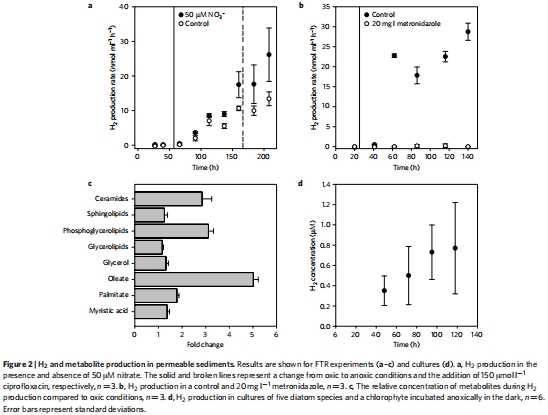
數據來源:圖1a, b; 圖2a, b。
抗生素與抑制劑實驗數據(識別主導生物域和特定途徑)
測量指標:在添加阿莫西林(抗生素)和 甲硝唑(特異性抑制劑)后,DIC和H?產生速率的變化。
研究意義:圖1d顯示,阿莫西林處理對DIC生產無顯著影響,而圖2b顯示甲硝唑完全抑制了H?生產。這構成了論文最關鍵的證據鏈:DIC/H?生產不依賴于細菌(耐抗生素),但依賴于真核生物的特定發酵途徑(對甲硝唑敏感)。這直接將主導代謝者鎖定為真核微藻,并將代謝途徑鎖定為鐵氧還蛋白/氫化酶介導的暗發酵。
數據來源:圖1d; 圖2b。
代謝組學數據(揭示細胞內代謝產物變化)
測量指標:厭氧與有氧條件下,沉積物中脂質(如磷脂酰甘油酯、神經酰胺)和特定脂肪酸(如油酸)的相對濃度變化。
研究意義:圖2c顯示,厭氧條件下脂質含量顯著增加(3-5倍)。這為暗發酵途徑提供了分子水平的證據,與微藻在發酵過程中將丙酮酸/乙酰輔酶A轉化為儲能脂質的理論預測完全一致,解釋了碳的流向。
數據來源:圖2c。
純培養微藻產H?數據(提供最直接的驗證)
測量指標:從沉積物中分離的多種硅藻(Fragilariopsis sp.) 和一種綠藻(Pyramimonas sp.)在黑暗厭氧培養中產生的H?濃度。
研究意義:圖2d清晰表明,所有測試的微藻純培養物在厭氧條件下均能產生H?。這是支撐“微藻是沉積物中H?來源”這一假說的決定性證據,排除了沉積物復雜群落中其他生物的干擾,使因果關系非常明確。
數據來源:圖2d。
細胞計數與生物量估算(確認微藻的優勢地位)
測量指標:沉積物中微藻的細胞豐度和分類組成,以及基于葉綠素a含量估算的微藻生物量碳。
研究意義:正文中指出,硅藻等微藻非常豐富(>10? cells mL?1),其生物量碳約占沉積物總有機碳的57%。這為“微藻是主要代謝者”的論點提供了生物量基礎,說明它們有足夠的體量來驅動觀測到的宏觀代謝通量。
數據來源:正文中“細胞計數”部分及補充表2。
研究結論
真核微藻的暗發酵主導碳礦化:本研究提供了強有力的證據,證明在透水性沉積物的厭氧環境中,真核微藻(尤其是硅藻)的暗發酵是溶解無機碳(DIC)生產的主要途徑,其貢獻超過了細菌的硝酸鹽還原、硫酸鹽還原等傳統厭氧呼吸作用。
發酵是微藻的關鍵生存策略:微藻通過暗發酵來降解內部儲存的有機物(如淀粉),產生少量的ATP、H?、CO?,并將碳流向合成儲能分子(如脂質),從而在動態沉積物環境造成的黑暗、缺氧條件下長期存活(數周)。這解釋了它們在這種惡劣環境中的持久性。
H?是發酵的關鍵指示物:觀測到的高濃度H?直接源于微藻的鐵氧還蛋白/氫化酶途徑。H?的釋放表明發酵過程正在活躍進行,并可能為沉積物中的氫營養型細菌提供底物,從而連接了真核生物和原核生物的代謝網絡。
挑戰傳統范式:本研究挑戰了基于氧化還原級聯(Redox Cascade)的傳統沉積物厭氧代謝模型。在富含微藻的透水沉積物中,基于真核生物的發酵可以成為比基于細菌的終端電子受體呼吸更重要的碳循環通道。
具有全球意義:考慮到透水性沉積物和底棲微藻在全球海岸帶的廣泛分布,微藻暗發酵可能是一個重要但被忽視的全球性碳礦化和能量流動途徑,對全球生物地球化學模型具有重要啟示。
丹麥Unisense電極測量數據的研究意義
在本研究中,丹麥Unisense H?微電極是發現和定量化暗發酵過程不可或缺的關鍵工具,其研究意義至關重要:
實現了對H?的高分辨率、實時原位測量:Unisense H?微電極能夠連續、高頻率地測量流通式反應器(FTR)出口水中溶解H?的濃度。這種原位、實時的測量能力至關重要,因為H?是一種溶解性低、周轉速率快的氣體,傳統的氣相色譜取樣方法可能會因時間延遲和樣品處理引入誤差,無法捕捉其動態變化過程。電極的高靈敏度使得檢測nM級別的H?變化成為可能,從而首次在環境沉積物系統中清晰記錄到如此高強度的H?通量。
提供了暗發酵的“吸煙槍”證據:H?的產生是發酵過程的典型特征。Unisense電極測量到的H?數據(圖2a)是推斷暗發酵存在的直接實驗證據。數據顯示H?在厭氧開始后數十小時才出現,并持續增長,這反映了微生物群落從有氧代謝向厭氧代謝的轉換和發酵過程的啟動。沒有這種直接的H?通量測量,暗發酵的假設將缺乏最有力的支持。
支撐了關鍵的藥理學實驗:Unisense電極在甲硝唑抑制實驗(圖2b)中發揮了決定性作用。通過連續監測H?濃度,研究者能夠清晰、定量地展示甲硝唑添加后,H?生產被迅速且完全地抑制。這一結果將產H?的代謝途徑特異性地指向了真核生物的[FeFe]-氫化酶系統,因為甲硝唑是該途徑的已知抑制劑。電極提供的實時響應數據使得這一因果關系非常明確。
驗證了純培養實驗的假說:在純培養微藻的驗證實驗中(圖2d),Unisense電極被用來確認分離出的硅藻和綠藻確實能夠在黑暗厭氧條件下產生H?。這一測量為“沉積物中的H?源自微藻”這一核心假說提供了最直接的、決定性的驗證,將宏觀通量與特定微生物群體的代謝功能直接聯系起來。
揭示了潛在的生態系統級相互作用:觀測到的凈H?釋放速率(盡管可能部分被細菌消耗)提示了沉積物中一個新的能量循環:微藻通過發酵產生H?,這些H?隨后可能被共生的氫營養型細菌(如硫酸鹽還原菌、產甲烷菌等)利用。Unisense電極測量的H?通量是量化這一潛在交叉喂養(cross-feeding)關系的基礎。沒有對H2的精確量化,就無法評估其在沉積物微生物食物網中的重要性。
綜上所述,丹麥Unisense H?微電極在本研究中扮演了“代謝過程探針”的角色。其獨特的高時間分辨率、原位測量能力,使其成為捕捉短暫且動態的H?信號、從而揭示整個暗發酵過程的關鍵。這項技術的應用,是成功論證“真核微藻暗發酵主導透水沉積物厭氧代謝”這一顛覆性結論的實驗基石。沒有它,本研究可能只會停留在DIC通量的測量,而錯過了指向發酵這一獨特機制的最關鍵線索——氫氣的產生。