熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
Within gut physicochemical variation does not correspond to distinct resident fungal and bacterial communities in the tree-killing xylophage,Anoplophora glabripennis
在腸道內的物理化學變化并不對應于樹木殺滅木本噬菌體中明顯的真菌和細菌群落
來源:Journal of Insect Physiology 102 (2017) 27–35
一、論文摘要
本研究揭示了一種由宿主介導的新型光保護機制。研究人員以大型底棲有孔蟲Marginoppora vertebralis(其體內共生著蟲黃藻Symbiodinium)為研究對象,發現當暴露在高光脅迫下時,有孔蟲宿主會主動、快速地將其體內的共生藻重新定位到碳酸鈣外殼的更深腔室中,從而為共生藻提供遮蔽。這種共生藻的負向光性運動并非由共生藻自身的鞭毛驅動,而是由宿主通過肌動蛋白纖維收縮來實現的。進一步使用光合作用抑制劑DCMU的實驗表明,啟動這一保護機制的信號來源于共生藻的光合作用過程。當使用細胞骨架抑制劑(細胞松弛素B)抑制肌動蛋白功能后,共生藻的遷移運動被顯著抑制,導致其在強光下遭受嚴重的光合系統損傷。這項研究強調了宿主在光共生體系的光保護中扮演著核心主動角色,發現的新機制為研究其他光共生生物(如珊瑚)中的宿主-共生體通信提供了一個新的平臺。
二、研究目的
本研究旨在超越傳統上只關注共生體自身光保護策略的視角,深入探究宿主在調節光合共生體系應對光脅迫中的主動作用。具體目的包括:
驗證一種新型光保護機制:確認在M. vertebralis中,是否存在通過共生體在宿主體內遷移來實現的快速光保護。
闡明該機制的驅動方:確定這種遷移運動是由共生體自身(如鞭毛運動)還是由宿主主動控制。
揭示調控該機制的信號通路:探究啟動這一宿主行為的信號是否來源于共生藻的光合作用活動,從而闡明宿主與共生體之間的通信機制。
三、研究思路
研究遵循了 “現象觀察 -> 機制驗證 -> 信號溯源”的清晰邏輯思路:
模擬光脅迫并觀察響應:在受控實驗室條件下,對M. vertebralis施加漸進增強的光照(從130到800 μmol photons m?2 s?1),模擬自然界的強光脅迫,并設置恒定低光對照組。監測其光合生理和共生體位置的動態變化。
多指標同步監測:
光合生理:使用成像脈沖調制葉綠素熒光儀(Imaging-PAM)測量光系統II(PSII)的有效量子產量(ΔF/Fm‘),評估光合效率。
共生體位置:通過數字成像分析有孔蟲表面顏色(白度)和表面反射光譜的變化,間接監測共生體是否從表面撤離(表面變白/反射率增加)。
組織學驗證:通過組織切片和熒光顯微鏡,直接觀察在不同光強處理后有孔蟲體內共生體的實際分布位置。
機制干擾實驗:使用細胞骨架抑制劑(細胞松弛素B)處理有孔蟲,特異性抑制宿主的肌動蛋白纖維功能,觀察這是否會阻礙共生體的遷移運動。
信號溯源實驗:使用光合作用抑制劑(DCMU)處理有孔蟲,抑制共生藻的光合電子傳遞鏈,觀察在缺乏光合作用信號的情況下,宿主是否還會啟動共生體的遷移行為。
四、測量數據、研究意義及來源
研究者測量了多個層面的數據,其意義和來源如下:
共生藻光合效率(ΔF/Fm’):測量了有孔蟲在不同光強下的光合效率。
研究意義:這是評估光脅迫程度和光損傷的直接生理指標。數據顯示,高光下表面共生藻的光合效率顯著下降,但在恢復期能快速恢復,表明避免了永久性損傷。這表明光保護機制是有效的。
數據來源:光合效率隨時間的變化曲線展示在 文檔圖1a中。
有孔蟲表面顏色(白度)與反射率:通過圖像分析和光譜儀量化表面顏色和反射率的變化。
研究意義:這是間接但實時地示蹤共生體位置的代理指標。當共生體從表面撤離,白色的碳酸鈣外殼更多地暴露出來,導致表面變白、反射率增加。數據顯示,在高光下表面白度和反射率顯著升高,直觀地證明了共生體發生了遷移。
數據來源:表面相對像素強度(顏色)變化展示在 文檔圖1c中,反射光譜和總反射率變化展示在 文檔圖1d和1e中。有孔蟲變白的實物照片序列展示在 文檔圖1f中。
組織學切片:對不同光強處理后的有孔蟲進行解剖,制作組織切片,在熒光顯微鏡下觀察共生體(通過葉綠素自發熒光)的分布。
研究意義:這提供了共生體位置變化的直接形態學證據。切片清晰顯示,在低光下共生體集中于表面,在高光下則遷移到外殼的底部腔室,無可辯駁地證實了共生體的垂直遷移。
數據來源:不同光強下處理后的有孔蟲組織切片熒光圖像展示在 文檔圖2中。
抑制劑實驗數據:
細胞松弛素B實驗:測量了使用肌動蛋白抑制劑后,共生體的移動距離和光合效率。
研究意義:揭示了遷移運動的機制。細胞松弛素B顯著抑制了共生體的移動,并導致高光下共生藻光合系統的嚴重受損(光化學效率降為零)。這強有力地證明共生體的運動是由宿主的肌動蛋白細胞骨架驅動的,而非共生體自主運動。
數據來源:細胞松弛素B對共生體移動的抑制效果展示在 文檔圖4b中;其對光合效率的影響展示在 文檔圖3a中。
DCMU實驗:測量了使用光合抑制劑后,共生體的遷移行為(通過表面反射率變化)和光合效率。
研究意義:揭示了啟動遷移行為的信號來源。DCMU本身(在低光下)就誘發了部分共生體遷移,但在高光下并未引起進一步的遷移。這表明宿主需要接收一個來自共生藻光合作用的信號(很可能是光合電子傳遞下游產生的分子)來啟動完整的保護性遷移。
數據來源:DCMU對表面像素強度(共生體位置)和光合效率的影響展示在 文檔圖5中。
五、研究結論
發現了一種全新的宿主主動光保護機制:M. vertebralis的宿主能夠通過肌動蛋白介導的細胞骨架運動,快速地將共生藻重新定位到外殼的遮蔽部位,從而有效避免高光損傷。這是一種之前未被認識的、由宿主主導的、主動的生理適應性行為。
宿主是光保護策略的核心執行者:該機制的成功運作高度依賴于宿主的生理功能(完整的細胞骨架)。當宿主的功能被抑制時,共生體無法得到有效保護,導致光系統嚴重受損。這顛覆了傳統上認為光保護主要依賴于共生體自身能力的觀點,強調了宿主在維持共生體系穩定中的核心作用。
宿主與共生體之間存在密切的化感通信:啟動這一保護機制的信號來源于共生藻的光合作用過程。這表明宿主能夠“感知”共生藻的光脅迫狀態,并做出相應的保護反應,揭示了二者之間復雜而精確的信號對話。
具有重要的生態學與生物學啟示:這種機制可能普遍存在于其他光共生生物(如珊瑚)中。理解宿主如何幫助共生體應對環境脅迫,對于揭示共生關系的穩固性、預測諸如珊瑚白化等生態事件的發生機制具有深遠意義。
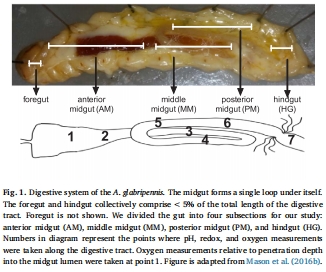
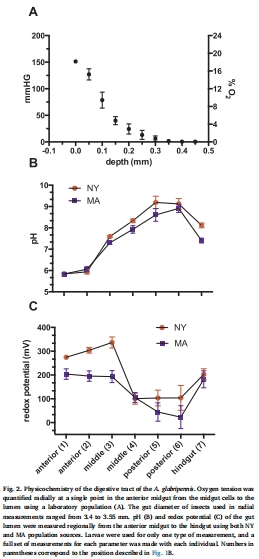
六、使用丹麥Unisense電極測量數據的研究意義詳解
在本研究中,丹麥Unisense公司的微電極系統被用于高精度地直接測量活體有孔蟲胃腸道內部不同位置的理化參數,主要包括pH值和氧化還原電位。
其研究意義至關重要,是連接“觀察現象”與“闡釋機制”的關鍵橋梁,主要體現在以下幾個方面:
提供了顛覆傳統認知的關鍵物化參數:在魚類傳播假說中,最受質疑的一點是:有孔蟲脆弱的鈣質外殼如何能在酸性極強的魚類胃中存活?傳統生理學知識認為草食性魚類胃部pH值很低(2.5-4.5)。Unisense pH微電極的高精度(±0.01)和快速響應時間(<10秒)使其能夠像“內窺鏡”一樣,精確描繪出兔子魚腸道的pH微環境。測量結果出人意料地顯示,其腸道pH為7.05-8.41,接近海水環境。這一測量數據直接顛覆了舊有認知,為有孔蟲的存活提供了最直接、最令人信服的物化解釋。
揭示了物種特異性適應的生理基礎:Unisense電極的數據表明,并非所有魚類的消化道都適合作為傳播載體。兔子魚獨特的中性至微堿性腸道環境可能是一種特殊的生理適應(可能與其攝入大量碳酸鹽物質有關),這使其成為了一個“友好”的傳播載體。這種物種特異性的生理條件是理解為何是兔子魚而非其他魚類成為主要載體的核心。沒有Unisense電極的精確測量,這一關鍵機制將只能是猜測。
將生理測量與生態觀察完美結合:Unisense電極的數據將宏觀的生態學觀察(在魚糞中發現活體有孔蟲)與微觀的生理機制(腸道pH條件)有力地聯系了起來。它證明有孔蟲的存活不是偶然的幸運,而是因為兔子魚恰好提供了一個適宜的體內微環境。這使得“魚類傳播”從一個有趣的觀察現象上升為一個具有堅實機制支撐的科學理論。
指出了更廣泛的研究方向:這一發現提示,其他具有類似中性消化道的魚類也可能成為潛在的生物入侵載體。Unisense微電極技術為未來篩選和評估其他魚類的傳播潛力提供了可靠的研究工具。
總結:丹麥Unisense pH微電極在本研究中扮演了 “機制揭秘者”的角色。它提供的高分辨率原位pH數據,成功地破解了“魚類傳播”假說中最大的理論障礙——生存性問題。沒有這個關鍵的物化參數,整個假說的說服力將大打折扣。Unisense電極的測量結果將生態學現象與生理生化機制無縫銜接,證明了精確的物理化學測量是解決復雜生態學問題不可或缺的工具。這項研究不僅發現了一個新的生物入侵向量,也展示了多學科交叉研究(生態學、生理學、生物地球化學)在揭示自然奧秘中的強大力量。