熱線:021-66110810,66110819
手機(jī):13564362870
熱線:021-66110810,66110819
手機(jī):13564362870
The Roles of Biofilm Conductivity and Donor Substrate Kinetics in a Mixed-Culture Biofilm Anode
混合培養(yǎng)生物膜陽極中生物膜電導(dǎo)率和供體底物動(dòng)力學(xué)的研究
來源:Environ. Sci. Technol. 2016, 50, 12799?1280
一、摘要概述
論文摘要指出,本研究通過實(shí)驗(yàn)評(píng)估了混合培養(yǎng)生物膜陽極中從供體底物(乙酸)到陽極的電子轉(zhuǎn)移(ET)動(dòng)力學(xué)和熱力學(xué)。研究采用了一個(gè)修改的生物膜傳導(dǎo)模型,該模型將電子轉(zhuǎn)移分為三個(gè)串聯(lián)步驟:(1)細(xì)胞內(nèi)電子轉(zhuǎn)移(IET),(2)從外膜蛋白到胞外輔因子(EC)的非歐姆細(xì)胞外電子轉(zhuǎn)移(EET),以及(3)通過生物膜基質(zhì)中的歐姆傳導(dǎo)從EC到陽極的電子轉(zhuǎn)移。在微型微生物電化學(xué)細(xì)胞(MxC)中,固定陽極電位為-0.15 V(相對(duì)于標(biāo)準(zhǔn)氫電極)時(shí),穩(wěn)態(tài)電流密度為0.82±0.03 A/m2。通過Illumina 16S rDNA和rRNA測序發(fā)現(xiàn),生物膜陽極中Geobacter屬的占比低于30%。生物膜導(dǎo)電性高達(dá)2.44±0.42 mS/cm,表明如果僅歐姆傳導(dǎo)EET受限,最大電流密度可達(dá)270 A/m2。歐姆傳導(dǎo)EET的能量損失可忽略不計(jì)(0.085 mV),第二步電子轉(zhuǎn)移的能量損失也較小(20 mV),而EC的電位為-0.15 V,表明超過99%的EC處于氧化狀態(tài)。乙酸利用的Monod動(dòng)力學(xué)相對(duì)較慢,至少87%的能量損失發(fā)生在細(xì)胞內(nèi)步驟。因此,對(duì)于高導(dǎo)電性生物膜,細(xì)胞內(nèi)電子轉(zhuǎn)移是電子從供體底物到陽極轉(zhuǎn)移的主要?jiǎng)恿W(xué)和熱力學(xué)瓶頸。
二、研究目的
本研究旨在定量評(píng)估混合培養(yǎng)生物膜陽極中電子轉(zhuǎn)移的速率限制步驟,特別是在高導(dǎo)電性生物膜背景下。通過結(jié)合實(shí)驗(yàn)測量和模型分析,識(shí)別電子轉(zhuǎn)移過程中的動(dòng)力學(xué)和熱力學(xué)瓶頸(如生物膜導(dǎo)電性、底物利用動(dòng)力學(xué)等),以優(yōu)化微生物電化學(xué)細(xì)胞(MxC)的性能。研究重點(diǎn)在于驗(yàn)證修改的Nernst-Monod模型(包括三個(gè)電子轉(zhuǎn)移步驟)的適用性,并闡明生物膜導(dǎo)電性與底物動(dòng)力學(xué)之間的相互作用。
三、研究思路
研究采用微型雙室微生物電化學(xué)細(xì)胞(MxC)作為實(shí)驗(yàn)平臺(tái),陽極由兩個(gè)金電極構(gòu)成,陰極為石墨板,使用陽離子交換膜分離。操作步驟如下:
MxC設(shè)計(jì)與操作:設(shè)計(jì)微型MxC(工作體積256 μL),固定陽極電位為-0.15 V,以乙酸為底物,在非底物限制條件下運(yùn)行。
電子轉(zhuǎn)移模型:提出一個(gè)三步驟電子轉(zhuǎn)移模型(圖1a),包括IET、非歐姆EET(從外膜蛋白到EC)和歐姆傳導(dǎo)EET(從EC到陽極)。模型基于修改的Nernst-Monod方程(方程4),納入了生物膜導(dǎo)電性(K_bio)的影響。
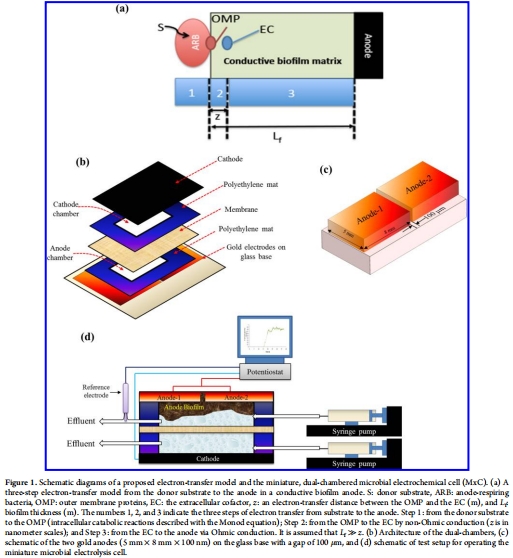
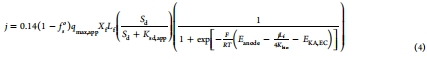
參數(shù)測量:在穩(wěn)態(tài)下,測量生物膜導(dǎo)電性、厚度、微生物群落結(jié)構(gòu)、循環(huán)伏安曲線(CV)和Monod動(dòng)力學(xué)參數(shù)。
數(shù)據(jù)分析:使用模型擬合實(shí)驗(yàn)數(shù)據(jù),計(jì)算各步驟的能量損失和動(dòng)力學(xué)參數(shù),識(shí)別瓶頸步驟。
研究思路強(qiáng)調(diào)通過多學(xué)科方法(電化學(xué)、微生物學(xué)、模型模擬)全面解析電子轉(zhuǎn)移過程。
四、測量數(shù)據(jù)及研究意義
以下列出關(guān)鍵測量數(shù)據(jù),注明來源(圖或表),并解釋其研究意義。數(shù)據(jù)來自文檔中的文本描述、表1、表2和圖2,避免使用表格形式,而是以描述性列表呈現(xiàn)。
穩(wěn)態(tài)電流密度(來自文本描述)
數(shù)據(jù):穩(wěn)態(tài)電流密度為0.82±0.03 A/m2。
研究意義:該電流密度相對(duì)較低,為后續(xù)分析電子轉(zhuǎn)移瓶頸提供基線。結(jié)合模型,表明電流輸出受IET限制而非EET。
生物膜厚度(L_f)使用丹麥Unisense電極測量(來自文本描述)
數(shù)據(jù):生物膜厚度平均為100 μm(通過Unisense微電極系統(tǒng)測量,重復(fù)測量值為85 μm和115 μm)。
研究意義:L_f是計(jì)算Monod動(dòng)力學(xué)參數(shù)和能量損失的關(guān)鍵變量。準(zhǔn)確測量厚度確保了模型參數(shù)的可靠性,并為計(jì)算歐姆傳導(dǎo)能量損失提供基礎(chǔ)。這部分將在第六部分詳細(xì)解讀。
生物膜導(dǎo)電性(K_bio)來自表2

數(shù)據(jù):K_bio為2.44±0.42 mS/cm(通過兩探針法測量,生物膜電導(dǎo)為0.63±0.11 mS,控制電導(dǎo)為0.37±0.00 mS)。
研究意義:高K_bio證實(shí)生物膜具有優(yōu)良的導(dǎo)電性,支持歐姆傳導(dǎo)機(jī)制。計(jì)算顯示EET步驟的能量損失極小(0.085 mV),表明EET不是電流限制因素。
微生物群落結(jié)構(gòu)來自表1

數(shù)據(jù):基于16S rDNA和rRNA測序,Geobacter屬在DNA庫中占27%,在RNA庫中占21%;其他優(yōu)勢菌包括Rhodopseudomonas屬(RNA庫16%)、Rhodocyclaceae科(接近Azoarcus和Thauera屬,RNA庫28%)等。
研究意義:群落多樣性(Geobacter占比低)解釋了較慢的Monod動(dòng)力學(xué),表明ARB種群結(jié)構(gòu)直接影響IET效率。高導(dǎo)電性盡管存在,但底物利用動(dòng)力學(xué)成為瓶頸。
循環(huán)伏安曲線(CV)和半飽和EC電位(E_KA,EC)來自圖2
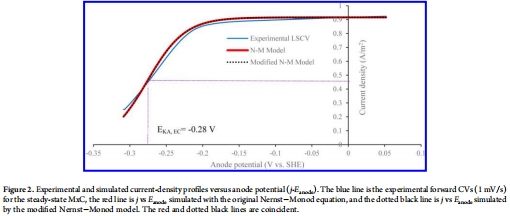
數(shù)據(jù):CV曲線呈S形,E_KA,EC為-0.28±0.00 V。
研究意義:E_KA,EC值較負(fù)表明電流密度在低陽極電位下飽和,有利于能量回收。CV與模型擬合良好(圖2),驗(yàn)證了修改的Nernst-Monod方程的準(zhǔn)確性,并支持EC以氧化態(tài)為主(>99%)。
Monod動(dòng)力學(xué)參數(shù)來自文本描述
數(shù)據(jù):表觀半飽和濃度(K_s,app)為274 g COD/m3,最大比底物利用率乘以活性生物膜密度(q_max,app X_f)為92×103 g COD/m3-d。
研究意義:K_s,app較高且q_max,app X_f較低,表明底物利用動(dòng)力學(xué)緩慢,這與Geobacter占比低一致。這些參數(shù)直接關(guān)聯(lián)IET步驟的能量損失(占總體87%),突出了IET的核心瓶頸作用。
五、結(jié)論
本研究得出以下結(jié)論:
細(xì)胞內(nèi)電子轉(zhuǎn)移(IET)是主要瓶頸:盡管生物膜導(dǎo)電性高(K_bio=2.44 mS/cm),但I(xiàn)ET步驟的能量損失占總能量損失的87%,且Monod動(dòng)力學(xué)參數(shù)(如q_max,app X_f)較低,導(dǎo)致電流密度受限。EET步驟的能量損失可忽略(0.085 mV),表明歐姆傳導(dǎo)效率高。
模型驗(yàn)證:修改的Nernst-Monod模型成功描述了電子轉(zhuǎn)移過程,CV數(shù)據(jù)擬合良好,證實(shí)了EC以氧化態(tài)為主(>99%),且非歐姆步驟(第二步)能量損失小(20 mV)。
應(yīng)用意義:對(duì)于高導(dǎo)電性生物膜,優(yōu)化MxC性能的關(guān)鍵在于提升IET動(dòng)力學(xué),如通過富集高效ARB(如Geobacter)或增加生物膜密度(X_f)。生物膜導(dǎo)電性本身不是限制因素,但需與底物動(dòng)力學(xué)協(xié)同優(yōu)化。
這些結(jié)論為設(shè)計(jì)高效微生物電化學(xué)系統(tǒng)提供了理論指導(dǎo),強(qiáng)調(diào)需平衡生物膜導(dǎo)電性和微生物代謝活性。
六、詳細(xì)解讀使用丹麥Unisense電極測量數(shù)據(jù)的研究意義
丹麥Unisense電極在本研究中用于精確測量生物膜厚度(L_f),其研究意義至關(guān)重要:
測量原理:Unisense微電極系統(tǒng)(型號(hào)MM33)連接電機(jī)驅(qū)動(dòng)微操縱器,使用不銹鋼導(dǎo)電微電極(針尖直徑100 μm)步進(jìn)移動(dòng)(步長5 μm)向生物膜表面接近。通過監(jiān)測電阻變化(從開路狀態(tài)到接觸生物膜外層時(shí)電阻降至MΩ級(jí),接觸陽極表面時(shí)降至<1 Ω)確定L_f。測量過程中,為避免脫水,在生物膜表面滴加底物培養(yǎng)基,并在20分鐘內(nèi)完成以減少氧氣影響。
研究意義:
數(shù)據(jù)準(zhǔn)確性保障:Unisense電極提供高空間分辨率(微米級(jí)),能準(zhǔn)確量化L_f(如平均100 μm)。這些數(shù)據(jù)是計(jì)算Monod動(dòng)力學(xué)參數(shù)(如q_max,app X_f)和能量損失的基礎(chǔ),確保了模型的可靠性。例如,L_f用于計(jì)算歐姆傳導(dǎo)能量損失(ΔE_EET = j L_f / 4 K_bio),結(jié)果為0.085 mV,證實(shí)EET步驟非限制性。
支持動(dòng)力學(xué)分析:L_f測量直接用于修改的Nernst-Monod方程(方程4),幫助區(qū)分IET和EET的貢獻(xiàn)。穩(wěn)定且均勻的L_f(重復(fù)測量偏差小)表明生物膜結(jié)構(gòu)完整,從而突出IET動(dòng)力學(xué)下降的主導(dǎo)作用,而非物理厚度變化。
技術(shù)優(yōu)勢:Unisense電極的非破壞性測量最小化了對(duì)生物膜的干擾,電流密度在測量后能迅速恢復(fù)穩(wěn)態(tài),體現(xiàn)了該方法在活體生物膜研究中的適用性。這為未來MxC研究提供了可靠的厚度監(jiān)測標(biāo)準(zhǔn),特別是在微型系統(tǒng)中。
總之,Unisense電極數(shù)據(jù)不僅驗(yàn)證了生物膜的物理特性,還強(qiáng)化了結(jié)論的嚴(yán)謹(jǐn)性——即電流限制主要源于IET動(dòng)力學(xué)而非生物膜結(jié)構(gòu)或EET。該測量方法為定量分析電子轉(zhuǎn)移瓶頸提供了關(guān)鍵技術(shù)支持。