熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
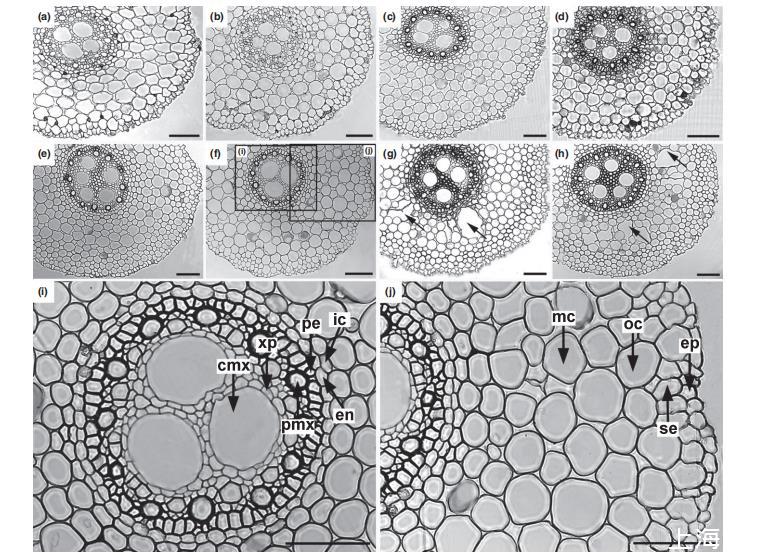
不定根不同細胞類型的徑向K、Na和Cl濃度
確定了在停滯溶液中的根,其根的中柱和尖端區(qū)域發(fā)生嚴重O?缺乏后,我們評估了這些組織中O?缺乏是否影響細胞特異性K、Na和Cl濃度。測量了不定根(圖2i,j)中不同細胞類型的K、Na和Cl的液泡濃度(參見結果部分;cf.Lauchli et al.,2008),采樣位置在根尖后方10 mm和50 mm處。測量徑向離子剖面的根解剖結構包括表皮、皮層和中柱(圖2i,j)。皮層包括亞表皮、六到八層皮層細胞和一個內皮層(圖2j)。中柱中研究的細胞包括周細胞、木質部薄壁細胞和木質部導管。木質部由幾個外周后生木質部導管(早期后生木質部)和3-6個中心后生木質部(晚期后生木質部)導管組成(圖2i)。
鉀-距根尖10 mm在通氣非鹽根中,表皮和皮層細胞的鉀濃度相似,平均為57±3.5 mmol kg?1等效鮮重(圖4a)。鉀在周細胞和木質部薄壁細胞中最高(平均135±6.8 mmol kg?1),并向木質部導管方向下降,在中心后生木質部導管中最低(37±4 mmol kg?1)。通氣含鹽根中細胞的K濃度與通氣非鹽根中的細胞沒有統(tǒng)計學差異(P=0.4)。停滯處理導致根內鉀剖面與通氣處理中的不同,在兩種停滯處理的根中,周細胞的鉀濃度顯著降低(P<0.0001)。在這些缺氧根中,K有從亞表皮穿過皮層增加的趨勢,盡管在周細胞中相對較低(45pm 3.5 mmol kg^{-1}),但在木質部薄壁細胞(127±7 mmol kg?1)和中心后生木質部導管(38±6 mmol kg?1)中,K與通氣根中的濃度一樣高。
鉀-距根尖50 mm在通氣非鹽根中,表皮、亞表皮、外皮層和中皮層的K濃度相似(平均36±3 mmol kg?1;圖4b)。鉀濃度隨后在內皮層和內皮層中增加,在周細胞中達到最高水平(152±10 mmol kg?1),并向木質部導管方向下降,在中心后生木質部中達到最低水平(29±3 mmol kg?1)。通氣含鹽根中相應的鉀剖面模式與非鹽根相似,但通氣含鹽根中周細胞、木質部薄壁細胞和外周后生木質部導管的鉀濃度平均比這些細胞在通氣非鹽根中高70%(P<0.05)。在兩種停滯處理的根中,K濃度在皮層中與通氣根相似,但在周細胞、木質部薄壁細胞和外周后生木質部導管中顯著降低(P<0.0001)。
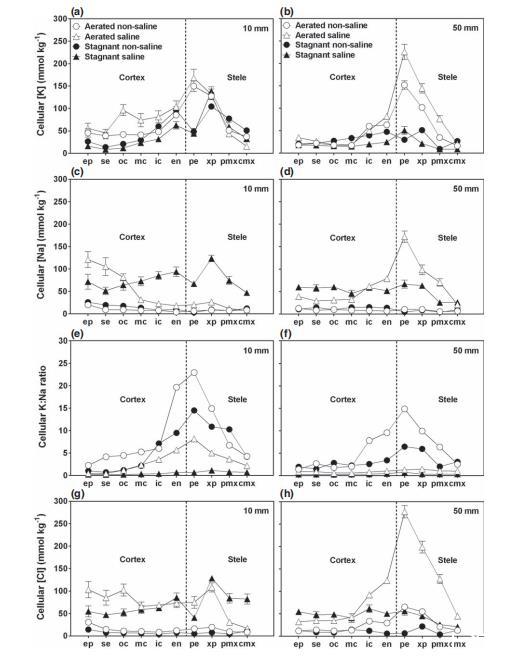
木質部鉀濃度及關于木質部成熟度的說明在所有處理中,距離根尖10 mm處的外周和中心后生木質部導管中的鉀濃度都相對較高,表明這些導管可能尚未成熟,含有細胞內容物而不是木質部汁液(圖4a)。這與Huang&Van Steveninck(1988)的研究一致,他們報道了在通氣營養(yǎng)液中生長的年輕大麥幼苗種子根距離根尖10 mm處,木質部導管中鉀濃度很高。在他們的研究中,距根尖50 mm處外周后生木質部的K濃度下降,但在距離根尖達150 mm處中心后生木質部的K濃度仍保持較高水平。對于本研究中的根,我們不確定距離根尖50 mm處的木質部導管是否成熟。在14-17天水培生長的大麥幼苗不定根(最大長度75 mm)中,軸向水力阻力從根尖開始隨距離增加而降低,直到距根尖約60 mm處,表明直到這個位置并非所有木質部導管都已成熟(Knipfer&Fricke,2011)。在本研究中,在距離根尖50 mm處,外周后生木質部的鉀濃度在通氣溶液中生長的根中往往高于在停滯溶液中生長的根(圖4b)。對于停滯處理中生長較慢的根,在距離根尖相同距離處的組織會比通氣處理中生長較快的根更老。因此,在停滯處理中,生長較慢的根,其木質部導管可能在更靠近根尖處成熟。
鈉-距根尖后方10 mm非鹽根中的鈉濃度在所有分析的細胞中都很低,無論是通氣還是停滯處理,均不超過12 mmol kg?1(圖4c)。在通氣含鹽處理的根中,鈉濃度在表皮、亞表皮和外皮層中最高(平均103±10 mmol kg?1),在中皮層向內的細胞和木質部導管中較低(平均20±1.5 mmol kg?1)(圖4c)。停滯含鹽根中相應的Na剖面與通氣含鹽根中的顯著不同(P<0.0001)。停滯含鹽根中表皮和亞表皮的Na濃度平均比通氣含鹽根中低47%,外皮層相似,但從中皮層向內到木質部導管的細胞中平均高300%(P<0.05)。
鈉-距根尖后方50 mm在非鹽根中,通氣處理和停滯處理的所有分析細胞中鈉濃度都很低,均不超過10 mmol kg?1(圖4d)。在通氣含鹽根中,表皮、亞表皮、外皮層和中皮層細胞中的Na含量低且相似(平均33±1 mmol kg?1),然后在內皮層和內皮層中急劇增加,在周細胞中達到最高濃度(172±13 mmol kg?1),隨后向木質部導管急劇下降,在中心后生木質部中降至最低濃度(21±3 mmol kg?1)。對于停滯含鹽處理中的根,直到木質部薄壁細胞的Na濃度在根內細胞中相似(平均56±2 mmol kg?1),但在木質部導管中降至26±0 mmol kg?1。
與通氣含鹽根在距根尖10 mm和50 mm處顯示出不同的Na剖面模式(P<0.0001;圖4c,d)相反,停滯含鹽根在距根尖10 mm和50 mm處的剖面沒有統(tǒng)計學差異(P=0.7;圖4c,d)。此外,在停滯含鹽根中,在兩個位置,Na濃度在根內分布更均勻,因此,對于停滯含鹽處理中的根,沒有表現(xiàn)出通氣根中描述的那種明顯的細胞類型差異。
鉀鈉比-距根尖后方10 mm K:Na比率在通氣非鹽根中最高,從表皮的2.2到周細胞的23不等(圖4e)。鹽度和停滯處理都導致K:Na比率降低:在通氣含鹽和停滯非鹽根的細胞中,比率(分別)從亞表皮的0.4和0.7到周細胞的8.2和14.5變化,
鉀鈉比-距根尖后方50 mm在通氣非鹽根中,從中皮層向外細胞的K:Na比率相對較低(平均值約2),在周細胞中達到最高值(15),并向木質部導管下降,在中心后生木質部中最低(2.5)。停滯非鹽根中,表皮和皮層的K:Na比率較低(平均2.3),在周細胞和木質部薄壁細胞中升至約6,在木質部導管中降至約2.5。通氣含鹽和停滯含鹽處理的根中,所有細胞的比率均較低(平均0.7)。
氯離子-距根尖后方10 mm非鹽根中的氯離子濃度較低(通氣根:14±0.7 mmol kg?1;停滯根:7±0.5 mmol kg?1)(圖4g)。通氣含鹽溶液中,所有細胞的Cl濃度升高至平均87pm 4,mmol,kg^{-1}(P<0.05),但木質部導管中仍較低(27±4 mmol kg?1)。停滯含鹽根的Cl濃度在表皮、亞表皮和外皮層比通氣含鹽根低約48%,但在其他細胞中相似;木質部導管的Cl濃度高出3倍(83±8 mmol kg?1)。通氣含鹽根中,表皮/亞表皮的高Cl濃度與高Na形成電荷平衡(圖4c),而Cl在整個根半徑保持較高水平,可能與內部細胞高K濃度相關(圖4a)。
氯離子-距根尖后方50 mm非鹽根中,表皮至中皮層細胞的Cl濃度較低(平均12±0.7 mmol kg?1;圖4h)。停滯非鹽根中Cl保持低位,而通氣非鹽根中Cl在內皮層和內皮層顯著增加,在周細胞中達峰值(65±3 mmol kg?1)。通氣含鹽根中,表皮至中皮層細胞Cl濃度相似(37±2 mmol kg?1),隨后在內皮層急劇上升,在周細胞中最高(278±14 mmol kg?1),最終在中心后生木質部降至45±3 mmol kg?1。停滯含鹽根中,Cl濃度在所有細胞中較均勻(平均50±2.5 mmol kg?1),在中心后生木質部為21±0.8 mmol kg?1。
關于蒸騰作用對木質部離子濃度影響的說明
X射線顯微分析樣品在實驗室采集(蒸騰速率低于溫室)。根尖區(qū)(10 mm)因木質部未成熟呈"水力隔離"狀態(tài)(Frensch&Steudle,1989),離子濃度受蒸騰影響小。距根尖50 mm處部分木質部或已成熟,蒸騰降低可能提高絕對離子濃度(Munns,1985),但處理間比較仍具參考價值。
不定根整體組織離子濃度
為補充細胞特異性數(shù)據(jù),測定了不定根整體離子濃度(圖5)。
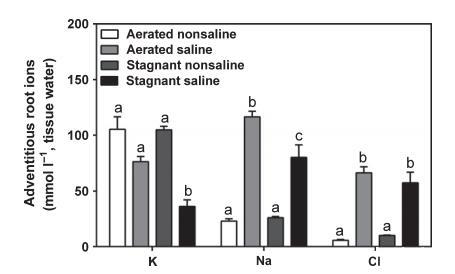
?K濃度:通氣非鹽、通氣含鹽和停滯非鹽處理相似(76-105 mmol L?1組織水);停滯含鹽處理降至對照的34%(P<0.05)(圖5)。
?Na濃度:通氣/停滯非鹽處理均低(22-26 mmol L?1);通氣含鹽和停滯含鹽處理分別比對照高5.1倍和3.5倍(P<0.05)。
?Cl濃度:非鹽處理中較低(6-10 mmol L?1);含鹽處理比對照高10-12倍(P<0.05)。
K:Na比率(表2):
通氣非鹽根最高(4.6);停滯含鹽根最低(0.4)。鹽度和缺氧均顯著降低該比率(P<0.05)。
表2大麥不定根K:Na比率
處理K:Na比率
通氣非鹽4.6±0.06a
通氣含鹽0.7±0.05b
停滯非鹽4.0±0.08c
停滯含鹽0.4±0.03b
(不同字母表示顯著差異,P<0.05)
討論
鹽度與缺氧顯著影響大麥根離子分布。微電極剖面(圖3)證實停滯溶液中根的中柱和根尖區(qū)O?缺乏最嚴重(孔隙度14-19%,表1)。缺氧降低K:Na比率(表2;圖4e,f),停滯含鹽根中該比率低至0.4(表2),細胞水平僅0.2-1.1(圖4e,f)。
關鍵發(fā)現(xiàn):
1.細胞特異性Na積累模式
?通氣含鹽根中,距根尖10 mm處Na富集于表皮/外皮層(圖4c),可能減少內部細胞Na暴露(此區(qū)域木質部未成熟,水流受限)。
?距根尖50 mm處Na在周細胞積累(圖4d),與硬粒小麥(Lauchli et al.,2008)和葡萄(Storey et al.,2003)一致,表明周細胞限制Na向木質部運輸。
?停滯含鹽根中Na分布均勻(圖4c,d),缺氧可能抑制液泡區(qū)室化(依賴H?-ATP酶供能)及質膜SOS1介導的Na?外排。
2.K裝載的細胞位點
通氣條件下,周細胞和木質部薄壁細胞均積累高K(圖4a,b),表明二者參與K向木質部裝載。缺氧降低周細胞K濃度(圖4b),證實該過程依賴能量。
3.徑向O?擴散屏障
微電極剖面(圖3)顯示表皮/中柱組織對O?擴散的高阻力導致pO?陡降。停滯溶液中,距根尖40 mm以遠區(qū)域中柱嚴重缺氧(pO?≤0.2 kPa),限制呼吸供能。
結論
中柱缺氧抑制周細胞和木質部薄壁細胞的能量依賴性K裝載。鹽脅迫下,周細胞對Na/Cl轉運的調控因缺氧受損。根尖區(qū)Na在外層細胞的積累(通氣條件)可減少內部細胞Na暴露,但該機制在缺氧時失效。
 相關新聞
相關新聞