熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
為了觀察細菌在雞糞甲烷發酵厭氧消化過程中的特定生態功能,選擇了豐度超過0.5%的24個操作分類單元,并與NCBI數據庫中的序列進行比較。基于在甲烷發酵過程中扮演的角色,它們被分為功能群。最豐富的群是碳水化合物降解和發酵細菌,包括15個屬,所有這些都有能力降解多糖如纖維素和淀粉,以及單糖如葡萄糖。Candidatus Cloacamonas sp.被分類為第二豐富群。Gallicola sp.,可以利用尿酸和肽生產揮發性脂肪酸,在兩個消化器中檢測到豐度為5%,這可能負責雞糞消化器中的高氮濃度。屬Fermentimonas、Dethiosulfatibacter、Peptoniphilus和Ignatzschineria也被鑒定為蛋白質降解細菌。
Syntrophaceticus schinkii作為乙酸代謝中的基本細菌,與氫營養型古菌形成共生關系,在兩個消化器中以低豐度0.04%檢測到。在中溫消化器中檢測到的互養乙酸氧化細菌包括Syntrophaceticus sp.、Tepidanaerobacter sp.和Clostridium sp.,它們通常與氫營養型產甲烷菌如Methanoculleus sp.、Methanosarcina sp.和Methanobrevibacter sp.共培養。
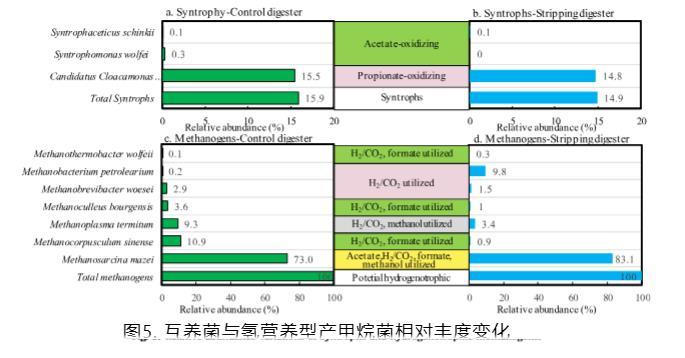
如圖5所示,產甲烷菌的組成和潛在的產甲烷途徑通過原位剝離發生變化。在屬水平上,Methanosarcina sp.在兩個消化器中均占主導。盡管Methanosarcina sp.有能力通過乙酸裂解產甲烷和SAO-HM從乙酸形成甲烷,但它主要被識別為乙酸裂解產甲烷菌。據報道,Methanosarcina sp.在高水平總銨態氮(5.8 g/L)、揮發性脂肪酸(2.6 g/L)和短hydraulic retention time(1.7天)時是優勢產甲烷菌。當進行原位氨氣剝離時,Methanosarcina sp.的相對豐度增加了10%。同時,氫營養型產甲烷菌的相對豐度,包括Methanocorpusculum sp.、Methanoplasma sp.、Methanoculleus sp.、Methanobrevibacter sp.、Methanobacterium sp.和Methanothermobacter sp.從27.0%降至17.0%。因此,提議原位氨氣剝離改善了產甲烷菌群落的結構,因為據報道乙酸裂解產甲烷菌在甲烷生產中比氫營養型產甲烷菌有優勢。
3.4.通過原位氨氣剝離增強產甲烷活性
在表3中,基于乙酸濃度,剝離消化器中的乙酸裂解特異性產甲烷活性比對照消化器高38%-294%。對照和剝離消化器中最大乙酸裂解特異性產甲烷活性分別為0.11和0.28 g-COD/(g-VSS·d),均在約10 g-COD/L乙酸時發現。當總銨態氮濃度從6.8降至5.8 g/L時,特異性產甲烷活性增加了156%。在先前研究中,報道了在總銨態氮2.4 g/L時雞糞消化的更高特異性產甲烷活性為0.56 g-COD/(g-VSS·d)。在剝離消化器中,總銨態氮和揮發性脂肪酸濃度仍處于高水平,特異性產甲烷活性未達到最大。因此,仍需要進行通過增強剝離強度降低總銨態氮濃度的進一步研究。氫營養型產甲烷菌的甲烷生產是厭氧消化過程中甲烷形成的另一個關鍵途徑,特別是在氨抑制過程中。因此,產甲烷性能也與氫營養型特異性產甲烷活性密切相關。溶解氫濃度已顯示與厭氧消化過程穩定性密切相關,并且降低溶解氫水平先前被報道在厭氧顆粒污泥處理過程中顯著降低揮發性脂肪酸水平。在本研究中,在溶解氫濃度500μmol/L時,H2/CO2的特異性產甲烷活性增強了33%(表3)。
剝離消化器中增強的產甲烷性能與更高的特異性產甲烷活性正相關。因此,推測本研究中增強的特異性產甲烷活性是由總銨態氮降低引起的。
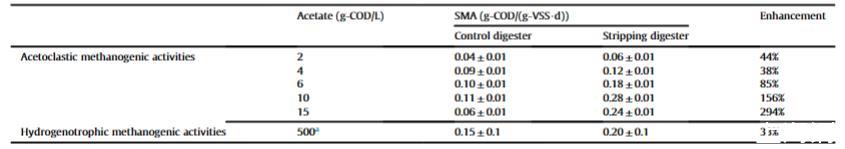
表3通過適度原位氨氣剝離增強產甲烷活性。
3.5.通過13C標記乙酸同位素示蹤量化的產甲烷途徑
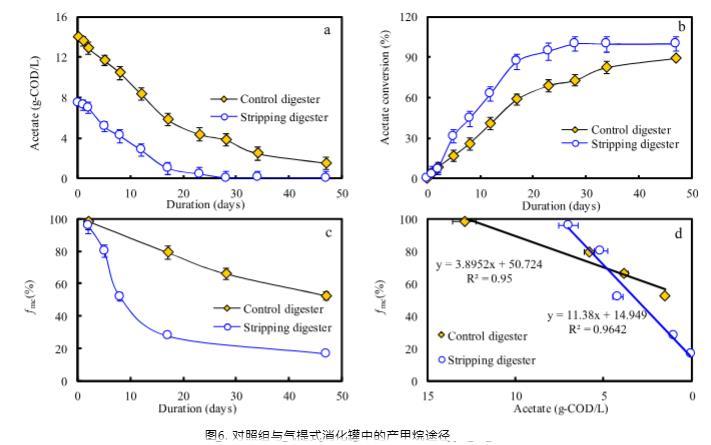
為了定量分析通過乙酸裂解產甲烷和互養乙酸氧化耦合氫營養型產甲烷途徑的產甲烷變化,進行了為期48天的批次測試(圖6)。在實驗期間,幾乎所有的乙酸都轉化為CH4(圖6a)。剝離消化器具有更快的乙酸轉化速率(圖6b)。通過H2還原CO2獲得的CH4比例,表示為f_mc,在對照和剝離消化器中分別從99%下降至96%-52%和16%(圖6c)。據報道,3 g/L的總銨態氮或6.0 g/L的乙酸高水平會誘導甲烷生產途徑從乙酸裂解產甲烷轉變為互養乙酸氧化耦合氫營養型產甲烷。在本研究中,消化器中存在高濃度的總銨態氮和乙酸。在乙酸降解過程中,通過線性擬合分析了f_mc與乙酸濃度之間的關系。剝離消化器中擬合出更高的斜率11.3,表明剝離消化器中乙酸濃度的變化可能更容易引起產甲烷途徑的轉變。
對照和剝離消化器中的平均f_mc分別為83%和77%,表明互養乙酸氧化耦合氫營養型產甲烷途徑在兩個消化器中均占主導地位。由氨氣剝離導致的較低總銨態氮有助于增加乙酸裂解產甲烷途徑的貢獻。
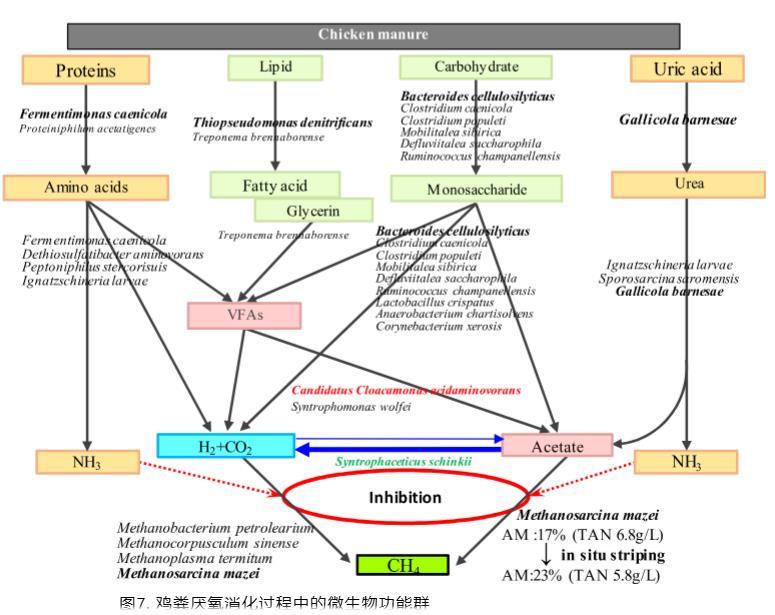
本研究中獲得的f_mc值是在較低總銨態氮水平2.4 g/L下雞糞消化所報道的f_mc值(41%)的兩倍。乙酸裂解產甲烷途徑被認為比互養乙酸氧化耦合氫營養型產甲烷途徑在乙酸降解和甲烷生產方面更有效。通過原位氨氣剝離降低總銨態氮濃度有助于降低剝離消化器中的f_mc值,這也是對互養菌和專性氫營養型產甲烷菌豐度降低的響應(圖5)。基于微生物群落和特異性產甲烷活性的分析,提出了雞糞高固體厭氧消化的代謝途徑模型,如圖7所示。
因此,較低的總銨態氮濃度以及產甲烷途徑從互養乙酸氧化耦合氫營養型產甲烷向乙酸裂解產甲烷的部分轉變,被認為是剝離消化器產甲烷性能增強的原因。值得注意的是,互養乙酸氧化耦合氫營養型產甲烷途徑的比例在原位氨氣剝離消化器中仍然非常高。如果原位氨氣剝離效率能夠顯著提高,則有可能進一步降低互養乙酸氧化耦合氫營養型產甲烷途徑的比例并增加甲烷產率。
4.結論
通過原位氨氣剝離,雞糞高固體厭氧消化的產甲烷性能在甲烷產率和揮發性脂肪酸去除方面均得到增強。由于總銨態氮濃度的降低,乙酸裂解和氫營養型產甲烷活性均得到增強。當甲烷八疊球菌屬起主要作用時,產甲烷途徑略微從互養乙酸氧化耦合氫營養型產甲烷向乙酸裂解產甲烷轉變。在雞糞高固體含量厭氧消化過程中,原位氨氣剝離技術部分緩解了氨抑制;然而,揮發性脂肪酸仍處于高濃度。因此,需要進一步研究以更大程度地加強氨氣剝離方法(例如提高氣流速率或頻率)。
 相關新聞
相關新聞