熱線:021-66110810,66110819
手機(jī):13564362870
熱線:021-66110810,66110819
手機(jī):13564362870
NsrR調(diào)控的M.catarrhalis基因的鑒定
DNA微陣列分析用于鑒定在全部三個(gè)nsrR突變體中,相對(duì)于相應(yīng)野生型親本菌株表達(dá)水平至少上調(diào)兩倍的基因。微陣列分析顯示,四個(gè)ORFs的表達(dá)水平在全部三個(gè)nsrR突變體中均一致增加至少兩倍(P<0.002)(圖5A)。這些ORFs是:norB(表達(dá)增加約30倍)、aniA(表達(dá)增加約30倍)、MC ORF 683(表達(dá)增加約10倍)和MC ORF 1550(表達(dá)增加2倍)。MC ORF 683編碼一種與來自Pseudoalteromonas haloplanktis的假設(shè)蛋白有39%同一性的蛋白。MC ORF 1550編碼一種與來自Psychrobacter species的光合反應(yīng)中心(PRC)桶狀結(jié)構(gòu)域蛋白有67%同一性的蛋白。PRC桶狀結(jié)構(gòu)域是在PRC亞基和一些參與RNA代謝的蛋白質(zhì)中發(fā)現(xiàn)的一個(gè)保守結(jié)構(gòu)域。進(jìn)行qRT-PCR分析以確認(rèn)這四個(gè)ORFs的表達(dá)受NsrR負(fù)調(diào)控。在M.catarrhalis O35E nsrR突變體中,所有四個(gè)ORFs的表達(dá)相較于野生型O35E親本菌株均上調(diào)(圖5B)。最后,一個(gè)推定的NsrR結(jié)合位點(diǎn)存在于所有這四個(gè)ORFs的上游區(qū)域。
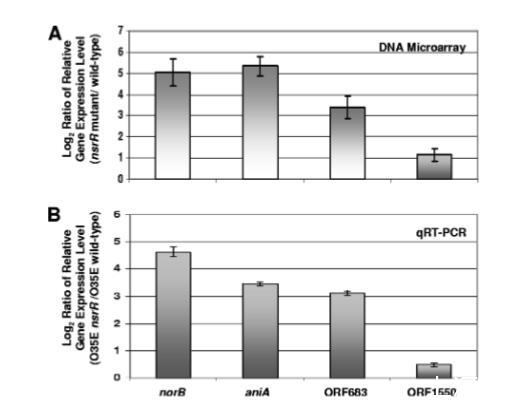
圖5.NsrR調(diào)控的卡他莫拉菌基因鑒定。(A)對(duì)野生型菌株O35E、7169和ETSU-9及其nsrR突變株提取的總RNA進(jìn)行DNA微陣列分析,方法詳見材料與方法部分。圖中顯示在所有三種nsrR突變株中持續(xù)上調(diào)至少兩倍(P<0.002)的基因數(shù)據(jù)。誤差棒表示標(biāo)準(zhǔn)差。(B)菌株O35E的nsrR突變細(xì)胞與野生型細(xì)胞中norB、aniA、MC ORF 683和MC ORF 1550表達(dá)相對(duì)水平的qRT-PCR分析。qRT-PCR測(cè)量的內(nèi)參對(duì)照為MC ORF 1234(62)的表達(dá)。誤差棒指示最大和最小相對(duì)表達(dá)水平。
不同氮化合物對(duì)M.catarrhalis NsrR調(diào)控基因表達(dá)的影響
進(jìn)行qRT-PCR分析以檢測(cè)NO??和NO對(duì)aniA、norB、MC ORF 683和MC ORF 1550相對(duì)表達(dá)水平的影響。需要指出的是,用于檢測(cè)aniA轉(zhuǎn)錄本的引物對(duì)位于aniA ORF的5'端內(nèi)部,但在aniA突變體中缺失區(qū)域的上游。因此,在qRT-PCR中使用該引物對(duì)可以檢測(cè)到aniA突變體中截短的aniA轉(zhuǎn)錄本。
在野生型O35E菌株(圖6A)和O35E aniA突變體(圖6B)中,添加NO??使aniA的轉(zhuǎn)錄增加了約五倍。將NO??添加到野生型O35E菌株細(xì)胞中導(dǎo)致產(chǎn)生易于檢測(cè)水平的NO,而O35E aniA突變體在相同條件下產(chǎn)生很少或沒有可檢測(cè)的NO(圖7A)。這些結(jié)果表明,NsrR調(diào)控的aniA表達(dá)對(duì)NO??敏感。
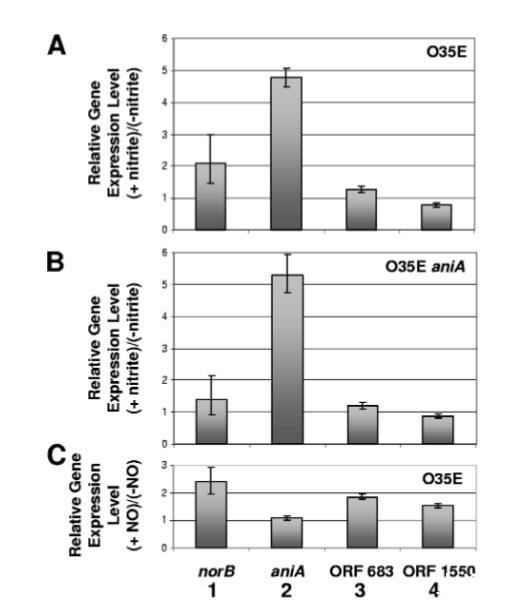
圖6.含氮化合物對(duì)NsrR調(diào)控基因表達(dá)的影響。(A和B)分別從在5 mM亞硝酸鈉存在或不存在條件下培養(yǎng)的卡他莫拉菌野生型菌株O35E(A)和O35E ani4突變體(B)細(xì)胞中提取的總RNA用于qRT-PCR。(C)從在NO生成化合物精胺NONOate存在或不存在條件下培養(yǎng)90分鐘的對(duì)數(shù)生長期卡他莫拉菌O35E細(xì)胞中提取的總RNA用于qRT-PCR。顯示了norB(條形1)、aniA(條形2)、MC ORF 683(條形3)和MC ORF 1550(條形4)的相對(duì)表達(dá)水平。qRT-PCR測(cè)量的內(nèi)參對(duì)照為MC ORF 1234的表達(dá)(62)。誤差條表示最大和最小相對(duì)表達(dá)水平。
添加NO??導(dǎo)致野生型O35E菌株中norB的表達(dá)上調(diào)(即兩倍)(圖6A),但未顯著增加O35E aniA突變體中norB的表達(dá)(圖6A)。推定的NsrR調(diào)節(jié)子中另外兩個(gè)ORFs(MC ORF 683和MC ORF 1550)的表達(dá)不受NO??存在的影響(圖6A和6B)。當(dāng)將NO生成劑spermine NONOate添加到M.catarrhalis野生型菌株O35E的對(duì)數(shù)期培養(yǎng)物中,90分鐘后norB的表達(dá)增加了約兩倍(圖6C)。相比之下,aniA的表達(dá)不受NO影響(圖6C)。這些結(jié)果直接證實(shí)了aniA的表達(dá)對(duì)NO不敏感,而norB的表達(dá)對(duì)NO敏感。
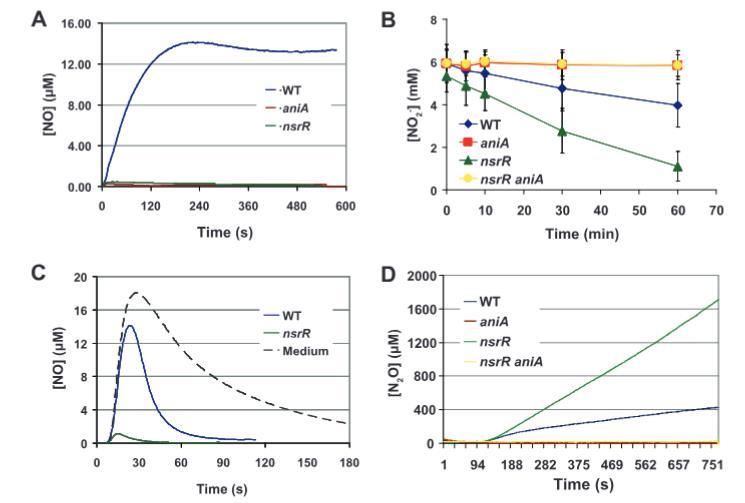
圖7.卡他莫拉菌野生型O35E及其突變株的含氮化合物代謝。(A)NO2-生成NO的過程。在補(bǔ)充5 mM亞硝酸鈉的BHI培養(yǎng)基中,使用一氧化氮特異性電極監(jiān)測(cè)野生型(WT)、aniA突變株和nsrR突變株的細(xì)胞懸液在10分鐘內(nèi)產(chǎn)生NO的情況。通過NO清除劑Carboxy-PTIO能消除可測(cè)量信號(hào)的能力證實(shí)了探針特異性(未顯示)。(B)NO2-的消耗。在補(bǔ)充5 mM亞硝酸鈉的BHI培養(yǎng)基中,采用格里斯反應(yīng)法監(jiān)測(cè)野生型、aniA突變株、nsrR突變株及nsrR aniA雙突變株細(xì)胞懸液對(duì)NO2-的消耗。(C)NO的消耗。將野生型和nsrR突變株的細(xì)胞懸液暴露于通過添加10 mM ProliNO產(chǎn)生的20μM NO環(huán)境中。使用一氧化氮特異性電極監(jiān)測(cè)溶解態(tài)NO濃度。作為參照,同時(shí)測(cè)定了BHI培養(yǎng)基的NO消耗活性(虛線所示)。(D)N2O的生成。在補(bǔ)充5 mM亞硝酸鈉(在檢測(cè)開始后2分鐘加入)的BHI培養(yǎng)基中,使用一氧化二氮特異性電極監(jiān)測(cè)野生型、aniA突變株、nsrR突變株及nsrR aniA雙突變株細(xì)胞懸液產(chǎn)生N2O的情況。
M.catarrhalis對(duì)氮化合物的代謝
野生型O35E菌株能從NO??產(chǎn)生NO,而O35E aniA突變體則不能(圖7A),這證實(shí)了M.catarrhalis AniA蛋白確實(shí)是一種亞硝酸還原酶,能夠通過還原NO??產(chǎn)生NO。此外,研究表明O35E nsrR突變體消耗NO??的速度遠(yuǎn)快于野生型親本菌株O35E(圖7B)。這很可能是由于O35E nsrR突變體中AniA表達(dá)升高所致,因?yàn)镺35E aniA突變體在同一時(shí)間段內(nèi)不消耗NO??(圖7B)。更重要的是,在O35E nsrR突變體中失活aniA基因完全消除了NO??的消耗(圖7B)。
對(duì)M.catarrhalis菌株消耗NO的檢查表明,O35E nsrR突變體消耗NO的速度遠(yuǎn)快于O35E野生型菌株(圖7C)。這很可能是由于nsrR突變體中norB表達(dá)增加所致。nsrR突變體對(duì)NO的快速消耗解釋了該突變體從NO??產(chǎn)生的可檢測(cè)NO水平非常低的原因(圖7A)。
在初步實(shí)驗(yàn)中,M.catarrhalis O35E nsrR突變體產(chǎn)生的N?O水平遠(yuǎn)高于其親本菌株;具體而言,nsrR突變體在大約50分鐘內(nèi)從5 mM NaNO?產(chǎn)生了2.5 mM N?O,而野生型親本菌株在相同時(shí)間內(nèi)僅產(chǎn)生約0.4 mM N?O。正如預(yù)期,失活aniA基因?qū)е翺35E aniA突變體和O35EΔnsrR aniA突變體均無法從亞硝酸鹽檢測(cè)到N?O的產(chǎn)生(圖7D)。
 相關(guān)新聞
相關(guān)新聞