熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
2.6.Acd_DPN7與FADH2的活性
我們測試了當FAD輔因子被還原為FADH2時,Acd_DPN7是否仍能催化3SP-CoA的脫亞磺酰。為了避免FADH2被氧氣再氧化為FAD,所有程序和測量均在含有福爾馬林氣體[N2:H2,95:5%(v:v)]氣氛的厭氧室中進行。該室配備有氧氣和氫氣分析儀,以確保在0 ppm的氧氣濃度下運行。在將所有溶液轉移到厭氧室之前,通過用氮氣吹掃除去氧氣。唯一的例外是蛋白質溶液,其中的氧氣如前所述被除去。用50 mM Tris-HCl緩沖液pH 7.4中的10 mM連二亞硫酸鈉滴定Acd_DPN7,將輔因子還原為FADH2。測定溶液含有終濃度為0.2 mM DTNB和0.2 mM 3SP-CoA,總體積為1 ml 50 mM Tris-HCl pH 7.4。溫度保持在7°C。通過加入10μl酶溶液開始測定。在412 nm處跟蹤吸光度的增加10分鐘。
2.7.天然Acd_DPN7及其與琥珀酰輔酶A復合物的結晶和晶體安裝
純化的Acd_DPN7最初通過氣相擴散在坐滴構型中結晶,蛋白質濃度為13 mg ml-1,使用The Classics II Suite條件D10[0.1 M Bis-tris pH 6.5,20%(w/v)聚乙二醇單甲醚5000],將400 nl蛋白質溶液與400 nl母液混合。晶體進一步優化,用于X射線衍射研究的晶體在24孔Linbro板中通過氣相擴散在懸滴構型中生長,來自0.1 M Bis-tris pH 6.5,5-20%PEG 3350,蛋白質濃度為10 mg ml-1,將2μl蛋白質溶液與2μl母液混合。天然Acd_DPN7晶體通過將晶體浸入含有10%甘油、0.1 M Bis-tris緩沖液pH 6.5、20%PEG 3350的溶液中進行冷凍保護。為了獲得Acd_DPN7與底物類似物的復合物,首先在存在4 mM琥珀酰輔酶A的情況下結晶Acd_DPN7蛋白質,然后在冷卻至100 K之前,將其浸泡在含有10 mM琥珀酰輔酶A以及冷凍保護緩沖液(0.1 M Bis-tris pH 6.5,30%PEG 3350,無甘油)的溶液中。
2.8.天然Acd_DPN7和浸泡琥珀酰輔酶A的Acd_DPN7的數據收集和處理
晶體學數據是使用同步輻射在PETRA III儲存環的EMBL P13光束線上收集的。晶體在100 K下用冷氮氣流冷卻。數據用XDS和AIMLESS處理。酶的脫輔基形式晶體衍射至1.89?分辨率,屬于空間群P 2?2?2,不對稱單元中有兩個單體,晶胞參數為a=75.6,b=100.6,c=118.5?。
該結構是使用自動晶體結構測定流程Auto-Rickshaw通過分子置換解決的,使用BALBES,以來自Thermus thermophilus HB8的酰基輔酶A脫氫酶晶體結構作為搜索模型。該結構使用ARP/wARP v.7.1、Coot和PHENIX精修至R因子和Rfree(5%數據)分別為16.2和20.1%。數據收集和精修統計數據見表1。
與底物類似物的Acd_DPN7復合物晶體衍射至2.30?分辨率,屬于空間群P 2?2?2,不對稱單元中有六個單體,晶胞參數為a=100.0,b=233.4,c=121.2?。該結構是通過使用Phaser進行分子置換解決的,以天然Acd_DPN7的單體A作為搜索模型。使用Coot和PHENIX進行精修,R因子和Rfree(5%數據)分別為19.9和24.4%。數據收集和精修統計數據見表1。
3.結果與討論
3.1.脫輔基Acd_DPN7的晶體結構
來自A.mimigardefordensis的3-亞磺酰丙酰輔酶A(3SP-CoA)脫亞磺酰酶(Acd_DPN7)的晶體結構是通過分子置換解決的,使用來自T.thermophilus HB8的酰基輔酶A脫氫酶(Acd)單體作為搜索模型,該酶具有36%的序列同一性。
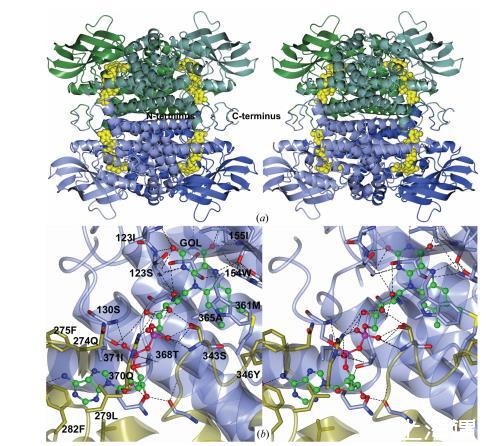
圖2(a)展示了來自A. mimigardefordensis菌株DPN7T的3-亞磺酰丙酰輔酶A(3SP-CoA)脫亞磺酸酶的立體結構。構成四聚體的兩個二聚體分別以綠色和藍色標示,二聚體的單體則用相應深淺顏色表示。FAD分子采用黃色空間填充模型呈現。(b)FAD分子(球棍模型:碳原子綠色、氧原子紅色、氮原子藍色)在A(淺藍色)和B(金色)兩個單體中共享。氨基酸殘基和甘油分子(GOL)通過粗鍵以淺藍色(單體A)或金色(單體B)標示,其中氧原子用紅色表示,氮原子用藍色表示。
尺寸排阻色譜證實了約150-200 kDa的天然分子量,如前所述確定,對應于觀察到的四聚體結構(圖2a)。使用PISA計算的埋藏表面積和自由解離能對于二聚體分別為7583?2和29.2 kcal mol?1,對于四聚體分別為23211?2和58.7 kcal mol?1。
Acd_DPN7的整體折疊與先前確定的其他Acds相似。單體由三個區域組成:一個初始的N端α-螺旋結構域(螺旋A-F)、一個β-折疊結構域(β-鏈1-7)和一個第二個C端α-螺旋結構域(螺旋G-K)。Acd_DPN7單體A和B的Cα原子與來自T.thermophilus HB8的Acd單體A的r.m.s.偏差分別為1.4和1.3?。這些r.m.s.d.值與其他Acds之間先前報道的值相當。
Acd_DPN7的幾個氨基酸殘基在整個Acd超家族中是保守的,并參與FAD結合,即K-X-W/F-I-T基序和GXXG基序。
每個FAD(圖2b)都位于β-鏈1的中間和β-鏈3的末端附近,腺嘌呤基團位于第二個單體的α-螺旋G和H之間。黃素基團與Ser124(A)和Thr156(A)的側鏈以及Ile121(A)、Ile123(A)、Ser124(A)、Trp154(A)和Thr156(A)的主鏈和兩個水分子形成擴展的氫鍵網絡。這兩個區域對應于通常在其他Acds中發現的TEPXXGS和KXW/FIT序列基序。FAD異咯嗪環的二甲基苯部分夾在Trp154(A)的一側和Ala365(A)的另一側之間,使口袋更具疏水性。在兩個單體的活性位點中,FAD異咯嗪環上方存在一個甘油分子(圖2b)。
核糖醇基團通過氫鍵與Ser343(B)的側鏈(GXXG基序的一部分)和幾個水分子相連。焦磷酸部分與Ser130(A)和Arg272(B)的側鏈以及局部水分子相互作用。腺嘌呤基團與Thr368(A)、Gln370(A)、Gln387(A)和Gln339(B)的側鏈形成氫鍵。存在幾個疏水殘基,如Phe275(B)、Phe282(B)、Leu285(B)和Leu279(B)。
使用PISA計算的二聚體的自由解離能為29.2 kcal mol?1,而去除兩個FAD分子的二聚體的自由解離能為17.6 kcal mol?1,這賦予了FAD在同源二聚體結構中的結構作用。
 相關新聞
相關新聞