熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
甲醛的轉化似乎由四氫葉酸依賴途徑或直接由甲醛脫氫酶介導(活性每分鐘每毫克蛋白質110 nmol)。甲醇脫氫酶活性為每分鐘每毫克蛋白質60 nmol,mxaF基因與Methylococcus capsulatus的mxaF有50%同一性。未發現sMMO的任何亞基。然而,鑒定出兩個完整的pmoCAB操縱子和一個具有部分pmoC的pmoCAB簇。發現與pmoA引物A189/A682有幾個(二至九個)錯配,解釋了從環境DNA中PCR擴增回收率低的原因。然而,PmoA的所有特征氨基酸都存在,而AmoA的特征氨基酸缺失。在所有細菌PmoA/AmoA蛋白中所有42個高度保守的氨基酸中,6至8個不被SolV菌株的一個或多個pmoA基因共享pmoA基因的系統發育分析顯示pmoA1和pmoA2與來自Solfatara和富集培養物的環境序列高度相似(圖1)。
pmoA3基因代表另一個全新的深分支。這三個新的pmoA序列一起表明,甲烷氧化細菌在系統發育上比目前假設的更為多樣化。最近的基因組數據顯示,一個Alpha或Gamma變形菌門甲烷氧化菌中可以存在兩個相同或遠緣相關的pmoA基因。使用特異性引物對甲烷生長的SolV細胞提取的mRNA進行RT-PCR證實了pmoA1和pmoA2信使RNA的表達。通過透射電子顯微鏡在SolV中未觀察到表達pMMO的甲烷氧化菌特征性的堆疊膜結構。相反,在用戊二醛固定或冷凍固定后觀察到約50-70 nm的圓形體。這些體可能讓人聯想到在嗜酸甲烷氧化菌Methylocella palustris中觀察到的囊泡。
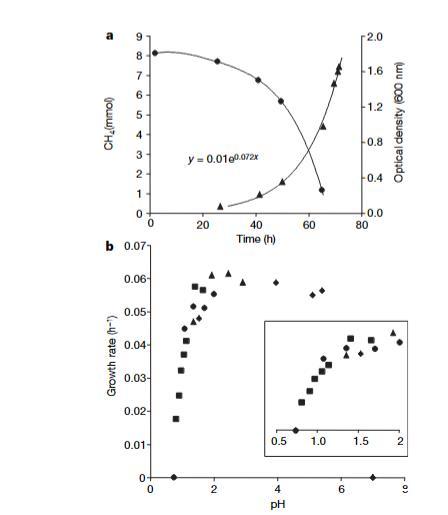
圖2|SolV菌株的生長特性。a,典型生長曲線顯示在pH 2和55°C條件下甲烷(圓形標記)的減少和光密度(三角形標記)的增加。該方程是通過數據點的最佳指數擬合。b,生長速率與pH值的關系。插圖展示了pH值低于2時的數據放大圖。不同符號表示在不同日期進行的實驗。
菌株SolV在pH 0.8至5.8之間生長(圖2)。最適溫度為55°C,在40°C以下和65°C以上僅觀察到輕微生長。甲烷上的最大比生長速率為0.07 h-1(倍增時間10小時)。二氧化碳和泥漿水的無機部分刺激生長。甲烷根據甲烷氧化菌的典型化學計量轉化為二氧化碳:CH4+1.6 O2→0.65 CO2+1.55 H2O+0.35 CH2O(生物量),每摩爾甲烷產率為6.4克干重。乙酸鹽、蘋果酸鹽、琥珀酸鹽、甲酸鹽、甲醛和酵母提取物(均為1 gl-1)在pH 2下完全抑制SolV在甲烷上的生長。該細菌顯然在低pH值下對由小有機酸引起的解偶聯非常敏感,因為在pH 5時甲酸鹽(pKa 3.75)不抑制生長。在100 mM NaCl以上或含葡萄糖的培養基中不發生生長。除甲烷外,氫氣也被氧化。菌株SolV在甲醇上生長良好,但添加的甲醇完全抑制甲烷消耗。甲醇耗盡后,甲烷消耗和生長僅在4小時后開始。乙烷抑制生長,盡管它作為競爭底物與甲烷同時以幾乎相同的速率轉化。乙炔(0.1%v/v)立即引起甲烷消耗的完全抑制,這一觀察結果支持pMMO是主要的甲烷氧化系統。SolV可以使用銨和硝酸鹽作為氮源。沒有甲烷時在銨上不發生生長。未觀察到固氮和厭氧硝酸鹽依賴的甲醇氧化。
SolV對甲烷的典型Ks值為6μM。然而,對氧的親和力異常高(Ks 0.7μM),反映了在其自然棲息地中需要競爭氧氣,其中微生物耗氧和持續流動的缺氧噴氣孔氣體(主要含二氧化碳)將導致氧氣濃度非常低。
使用主要覆蓋疣微菌目的探針EUBIII對分離物進行熒光原位雜交(FISH)分析,顯示強烈的雜交信號。使用EUBI、EUBII或alpha(ALF968)、beta(BET42a)或gamma(GAM42a)變形菌門探針未獲得信號。疣微菌門樣身份通過從焦磷酸測序獲得的其16S rRNA基因序列確認。基于該序列設計了一個特異性探針(SolV830,見方法),并與探針EUBIII一起使用以確認SolV培養物的純度。來自指數生長期培養物的所有細胞顯示雙重雜交。SolV的16S rRNA序列的系統發育分析表明,該分離物代表疣微菌門內一個新細分中的第一個成員(圖3)。成對距離分析顯示與其他細分的成員同一性低于81%。
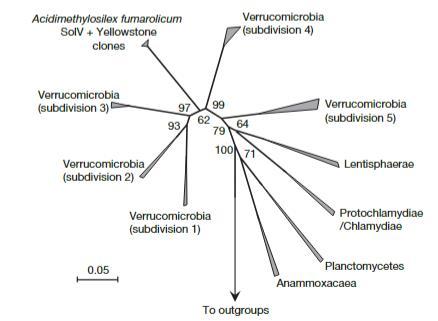
圖3|SolV菌株的16S rRNA基因序列與浮霉菌門(Planctomycetes)代表物種的系統發育關系
菌株SolV是首次報道的極端嗜酸甲烷氧化細菌,在系統發育上位于包含已知甲烷氧化菌的Alpha和Gammaproteobacteria亞門之外,我們建議將其命名為:“Acidimethylosilex fumarolicum”,新屬新種。
迄今為止,疣微菌門僅包含少數培養菌株,它們是厭氧或好氧異養菌,在或多或少復雜的培養基中生長于糖類上。然而,環境克隆文庫顯示疣微菌門存在巨大的生物多樣性,并且在許多生態系統(土壤、泥炭沼澤、酸性巖石排水和垃圾滲濾液)中經常以相對較高的數量出現,但生理學未知。有趣的是,推測廣泛分布的疣微菌門(其大多數成員尚未培養)可能與地球化學相關反應有關。使用菌株SolV 16S rRNA基因序列進行BLAST搜索顯示與六個環境克隆具有非常高的同一性(98-99%)(圖3),這些克隆是在黃石國家公園酸性熱泉(Rainbow和Joseph's Coat)微生物群落地球化學研究期間獲得的(未發表;NCBI登錄號:AY882698、AY882699、AY882710、AY882819、AY882820和AY882834)。這表明與A.fumarolicum相似的細菌可能是這些極端環境的常見居民。新的pmoA和16S rRNA基因序列可能有助于從較少極端的棲息地識別Planctomycetes/Verrucomicrobia/Chlamydiae超門甲烷氧化菌,并顯示它們的全球分布。
結論
該研究首次發現疣微菌門中存在極端嗜酸甲烷氧化功能,擴展了對甲烷氧化微生物多樣性的認知。菌株SolV對極端環境的適應機制(如高氧親和力、多pmoA基因)為其在酸性自然生境中的生態角色提供了依據,同時提示疣微菌門可能在全球碳循環中扮演尚未被重視的關鍵作用。
 相關新聞
相關新聞