熱線:021-66110810,66110819
手機(jī):13564362870
熱線:021-66110810,66110819
手機(jī):13564362870
對(duì)于肌紅蛋白的光譜掃描,將純化的馬肌紅蛋白蛋白在1毫克/毫升濃度下與1體積的10%抗壞血酸鈉混合以產(chǎn)生脫氧狀態(tài),然后進(jìn)行上述系列通氣以實(shí)現(xiàn)氧合肌紅蛋白狀態(tài)。
酵母的氧氣攝取。在過(guò)表達(dá)株和模擬株中,連續(xù)監(jiān)測(cè)了酵母懸浮培養(yǎng)物40分鐘內(nèi)的O2攝取速率。同時(shí)測(cè)量了以葡萄糖為碳源時(shí),酵母懸浮培養(yǎng)物從溶液中耗盡O2所需的時(shí)間。釀酒酵母菌株INVSc1用于表達(dá)HsAQP1、NtPIP1;3或AtPIP1;2,以及模擬對(duì)照株,按照上述方法培養(yǎng)并誘導(dǎo)異源蛋白表達(dá)。酵母在KH2PO4緩沖液(0.1M,pH 6)中洗滌兩次,然后懸浮于15mL經(jīng)N2鼓泡的SD-L-U+葡萄糖培養(yǎng)基中,置于50mL Falcon管中,使OD600達(dá)到5。向酵母懸浮液中通入空氣,直到其可溶性O(shè)2濃度達(dá)到約235μmol L-1,這是25°C下靜止培養(yǎng)基的穩(wěn)定飽和水平。從這一點(diǎn)開(kāi)始,使用連接到OXY-Meter(一種緊湊型O2微傳感器放大器,Unisense,Aarhus,丹麥)的尖端直徑為50μm的O2微傳感器(OX-50)每秒監(jiān)測(cè)并記錄酵母懸浮液中O2濃度的下降。使用Parafilm密封Falcon管,以盡量減少自由空氣擴(kuò)散。通過(guò)線性回歸計(jì)算初始0-1000秒內(nèi)O2濃度下降的斜率,其絕對(duì)值代表不同酵母株在相應(yīng)時(shí)間間隔內(nèi)的O2消耗速率。記錄每種酵母懸浮液的O2耗盡時(shí)間。均值和標(biāo)準(zhǔn)誤差基于六次生物學(xué)重復(fù)計(jì)算得出。
煙草根低氧研究:生長(zhǎng)條件與處理。煙草種子在土壤中萌發(fā),植物在受控環(huán)境生長(zhǎng)室中生長(zhǎng)兩周,維持22/18°C(晝/夜)溫度,60±10%相對(duì)濕度,以及18小時(shí)光周期,光合光子通量密度約為350微摩爾/平方米/秒。生長(zhǎng)兩周后,將植物轉(zhuǎn)移到含有半強(qiáng)度改良霍格蘭氏溶液的容器中,并用魚(yú)缸泵通氣。將36株植物隨機(jī)放置在六個(gè)容器中。一周后,通過(guò)用氮?dú)鉀_刷水使三個(gè)容器中的植物遭受低氧處理,達(dá)到溶解氧水平為2毫克/升,然后保持靜止。另外三個(gè)容器中的植物繼續(xù)通氣。
煙草中的定量RT-PCR。在低氧處理兩天和七天后取樣根和葉片。將組織樣品在液氮中使用研缽和研杵冷凍并勻漿。使用植物RNA提取試劑盒提取總RNA。如方法S1中所述進(jìn)行cDNA合成和qPCR。NtPIP1;1、PIP1;2、PIP1;3、PIP1;4和PIP2;1的轉(zhuǎn)錄豐度針對(duì)兩個(gè)參考基因EF1-α和核糖體蛋白L25的轉(zhuǎn)錄豐度的幾何平均值進(jìn)行標(biāo)準(zhǔn)化。使用引物設(shè)計(jì)軟件設(shè)計(jì)基因特異性引物。
ATP水平的測(cè)定。在低氧和通氣良好植物處理兩天和七天后,測(cè)量葉片以及根尖段、根中段和根基段的ATP水平。在處理兩天和七天后取樣煙草的根和葉片。將根分為基段、尖段和中段。將組織樣品研磨,并將50毫克研磨樣品置于2毫升離心管中的600微升冰鎮(zhèn)5%三氯乙酸中。將樣品劇烈渦旋20秒,冰上放置10分鐘,并在10,000倍重力、4°C下離心10分鐘。收集每份400微升上清液,并加入400微升冰鎮(zhèn)的Tris-乙酸緩沖液。對(duì)于ATP測(cè)定,將4微升混合物移液到96微升無(wú)ATP水中的96孔板的一個(gè)孔中。為了量化ATP,向每個(gè)孔中加入50微升來(lái)自ATP測(cè)定試劑盒的rLuciferase/Luciferin試劑,并按照制造商方案制備標(biāo)準(zhǔn)曲線。使用酶標(biāo)儀檢測(cè)生物發(fā)光信號(hào)。
統(tǒng)計(jì)分析。通過(guò)描述性統(tǒng)計(jì)計(jì)算每次測(cè)定中基于生物學(xué)重復(fù)的平均值和標(biāo)準(zhǔn)誤。使用單因素方差分析(Tukey檢驗(yàn))分析統(tǒng)計(jì)學(xué)差異。
結(jié)果
肌紅蛋白和水通道蛋白的蛋白質(zhì)表達(dá)及轉(zhuǎn)錄豐度。使用抗肌紅蛋白抗體的免疫印跡證明了在構(gòu)建的表達(dá)肌紅蛋白的選定酵母菌株中存在肌紅蛋白,但在INVSc1菌株中不存在。定量RT-PCR顯示,轉(zhuǎn)化酵母菌株中肌紅蛋白的轉(zhuǎn)錄豐度相似。使用抗人水通道蛋白1抗體的免疫印跡證明,該抗體能識(shí)別各自菌株中表達(dá)的異源水通道蛋白智人HsAQP1、煙草NtPIP1;3和擬南芥AtPIP1;2,并且也能微弱地識(shí)別僅表達(dá)肌紅蛋白的模擬菌株中的酵母同源水通道蛋白。比免疫印跡特異性更高的qRT-PCR檢測(cè)顯示,模擬菌株中異源表達(dá)的水通道蛋白基因HsAQP1、NtPIP1;3和AtPIP1;2的轉(zhuǎn)錄豐度可忽略不計(jì),但在每個(gè)相應(yīng)菌株中顯著較高。
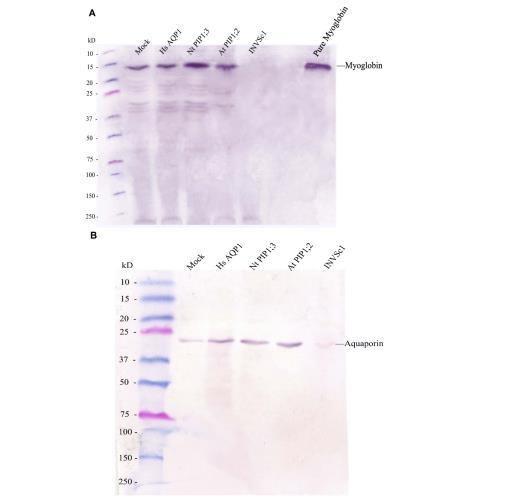
圖1.使用抗肌紅蛋白抗體和抗人水通道蛋白1抗體進(jìn)行免疫印跡分析。(A)使用一抗抗肌紅蛋白抗體對(duì)酵母總蛋白進(jìn)行免疫印跡。(B)使用一抗抗人水通道蛋白1抗體對(duì)酵母總蛋白進(jìn)行免疫印跡。
經(jīng)過(guò)甲醛固定、石蠟包埋以及制備用于抗人水通道蛋白1抗體免疫檢測(cè)的切片酵母細(xì)胞后,與相對(duì)較弱的細(xì)胞內(nèi)熒光信號(hào)相比,在HsAQP1菌株的酵母細(xì)胞切片外圍檢測(cè)到強(qiáng)烈的免疫熒光,表明質(zhì)膜可能是其定位位點(diǎn)。這與TargetP的亞細(xì)胞定位預(yù)測(cè)一致,提示不存在線粒體靶向肽,并指出分泌途徑是真核細(xì)胞中HsAQP1最可能的位置。
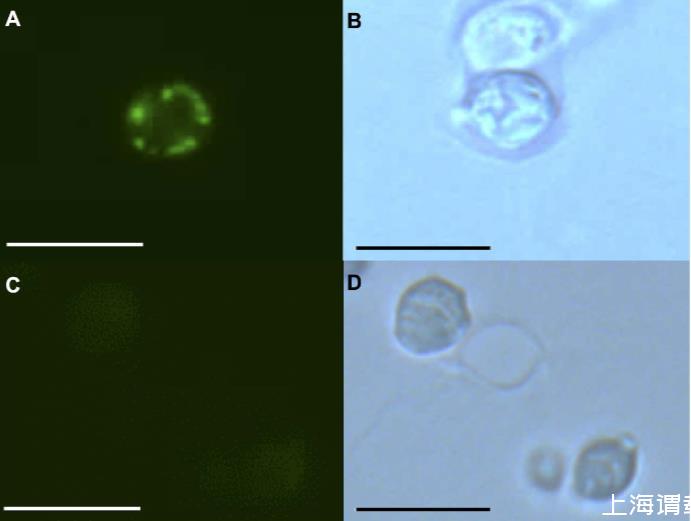
圖2.與一抗抗水通道蛋白1單克隆抗體和熒光素標(biāo)記的二抗孵育后,石蠟包埋的HsAQP1酵母菌株的間接免疫熒光結(jié)果。(A)藍(lán)光激發(fā)下的HsAQP1菌株。(B)明場(chǎng)下的HsAQP1菌株。(C)藍(lán)光激發(fā)下的模擬菌株。(D)明場(chǎng)下的模擬菌株。標(biāo)尺長(zhǎng)度為10μm。
氧氣轉(zhuǎn)運(yùn)。在所檢測(cè)的酵母菌株中,表達(dá)HsAQP1、NtPIP1;3、NtPIP1;4、NtPIP2;1和NtXIP1;1的菌株通過(guò)初步分光光度測(cè)量顯示出氧氣通透性統(tǒng)計(jì)學(xué)上的顯著增加,這表現(xiàn)為肌紅蛋白A541吸光度變化速率更高,與模擬對(duì)照相比。過(guò)表達(dá)擬南芥和雙色蠟?zāi)⒌乃ǖ赖鞍撞⑽锤淖傾541吸光度,表明對(duì)氧氣通透性沒(méi)有顯著影響。
在酵母原生質(zhì)體實(shí)驗(yàn)中,我們對(duì)兩種最具潛力的氧氣轉(zhuǎn)運(yùn)蛋白(HsAPQ1和NtPIP1;3)、一種在初步實(shí)驗(yàn)中未顯示氧氣轉(zhuǎn)運(yùn)特性的蛋白(AtPIP1;2)以及模擬菌株進(jìn)行了進(jìn)一步分析。基于對(duì)純化肌紅蛋白和酵母原生質(zhì)體的光譜掃描,我們選定ΔA541/ΔA600和ΔA319/ΔA341作為肌紅蛋白氧合指標(biāo)。90秒時(shí)的ΔA541/ΔA600值在各菌株中呈現(xiàn)與初步實(shí)驗(yàn)相同的趨勢(shì):表達(dá)NtPIP1;3的菌株數(shù)值最高,依次為HsAQP1、模擬菌株和AtPIP1;2(圖3)。經(jīng)過(guò)5次30秒曝氣處理5分鐘后,ΔA319/ΔA341值顯示出更顯著的統(tǒng)計(jì)學(xué)差異:HsAQP1與模擬菌株之間,以及所有表達(dá)肌紅蛋白的菌株與未轉(zhuǎn)化菌株INVSc1之間均存在明顯差異(圖4)。
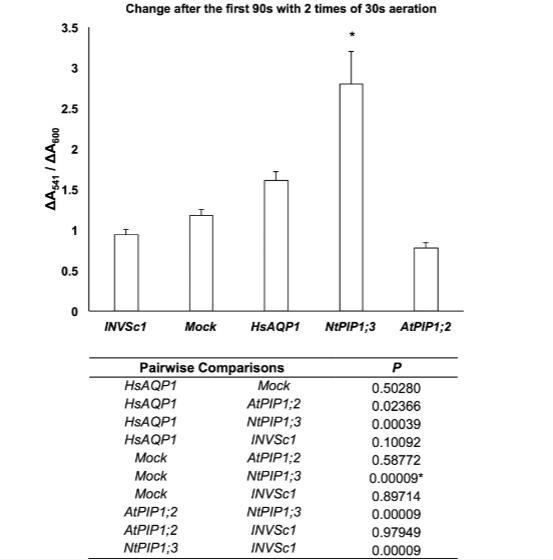
圖3.經(jīng)過(guò)2次30秒通氣后的前90秒內(nèi),酵母原生質(zhì)體的ΔA541/ΔA600值。星號(hào)表示與模擬菌株存在統(tǒng)計(jì)學(xué)顯著差異(下表顯示P值)(ANOVA,Tukey檢驗(yàn),P≤0.05,n=19±標(biāo)準(zhǔn)誤)。
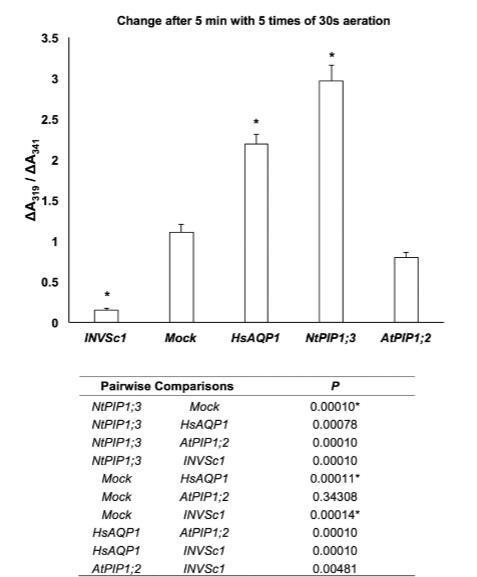
圖4.經(jīng)過(guò)5次30秒通氣共5分鐘后,酵母原生質(zhì)體的ΔA319/ΔA341值。星號(hào)表示與模擬菌株存在統(tǒng)計(jì)學(xué)顯著差異(下表顯示P值)(ANOVA,Tukey檢驗(yàn),P≤0.05,n=6±標(biāo)準(zhǔn)誤)。
 相關(guān)新聞
相關(guān)新聞