熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
每天早晚兩次檢查卵塊是否孵化,當培養皿中首次出現游泳的面盤幼蟲時,認為孵化開始,并立即用2%緩沖福爾馬林固定卵塊。之后將固定的卵塊在最厚區域切片,將切片平放在載玻片上。在100倍顯微鏡下,我們從邊緣到中心穿過中心進行直線樣帶,并對遇到的每個胚胎進行分期。分期鑒定為:擔輪幼蟲(光學顯微鏡下無區分特征,球狀)、前面盤幼蟲(形成明顯的面盤葉但無distinguishable的足或其他結構)、早期無殼面盤幼蟲(足、面盤和內臟團可區分但無殼)、早期有殼面盤幼蟲(淺色殼,暴露于福爾馬林后足和面盤未縮回)和晚期面盤幼蟲(完全形成的深色殼,動物完全縮入殼內)。每個樣帶中對17-36個胚胎進行分期。然后我們計算每個樣帶中有殼(晚期,孵化后可能存活)和無殼(早期,孵化后可能無法存活)胚胎的百分比。使用配對t檢驗分別檢查每個實驗中藻類對有殼面盤幼蟲比例的影響。
藻類對卵塊氧飽和度的影響
為了檢查卵塊中心氧飽和度如何隨藻類存在和不同光照條件變化,我們測量了上述實驗室實驗中2天齡和6天齡卵塊中心的氧氣。在兩天中,均在黑暗中等效8小時后和光照4小時后測量氧氣。為了確定實驗室測量是否與野外卵塊內部條件相似,我們還測量了低潮時單一潮池中卵塊的中心氧濃度,分別在白天和夜間進行。
本研究采用與克拉克式微電極(尖端直徑50微米,丹麥Unisense公司UWMeter型號)相連的水下便攜式皮安表,對野外和實驗室環境下的卵團氧氣濃度進行測量。該儀器探頭通過在空氣飽和水和氮氣飽和水中進行校準,校準溫度范圍覆蓋本研究所涉潮池的水溫變化區間。每個潮池均測量鹽度值,并相應調整氧飽和度計算參數。為評估潮池海水的氧合程度,我們在每個池內任意選取三個不同位點進行測量并取平均值。測量卵團內部氧氣濃度時,手動將探頭緩慢插入卵團最厚部位直至中心點,記錄該位點的氧氣測量值。每次完成測量后拔出探頭,重復兩次上述操作,每個卵團共獲得三個內部測量值。最終采用三個潮池測量值和三個內部測量值的平均數進行分析。
我們使用線性模型檢查在第2天和第6天藻類的存在是否影響卵塊中心氧飽和度,以及光照條件是否有影響。卵塊身份是模型中的隨機因子。數據分析前進行logit轉換,并滿足正態性和方差齊性假設。為了確定卵塊年齡是否影響中心胚胎的氧氣可用性,我們使用配對t檢驗比較每個卵塊在第2天與第6天(僅在藻類存在和光照條件下)測得的中心氧濃度。
潮池氧飽和度與卵塊氧飽和度的關系
為驗證潮池中的氧飽和度是否直接影響野外卵塊的氧飽和狀態,我們采用前述Unisense超微呼吸系統實時測量了潮池水體(n=6)與池中Siphonaria卵塊(每池n=2-10)的含氧量。氧濃度以每個潮池不同位點三次測量值的平均數計算。測量完成后,將卵塊采集至實驗室,在顯微鏡下根據觀察到的最新發育階段將其分為面盤幼蟲前期、早期面盤幼蟲期和晚期面盤幼蟲期。采用協方差分析(ANCOVA)檢驗水體氧飽和度(連續變量)與卵塊中心氧飽和度(因變量)的關系,并以發育階段作為分類因子。原始數據符合正態分布且方差齊性,無需進行轉換。
結果
藻類對發育時間和孵化的影響
實驗室實驗開始時,胚胎的發育階段從新產下(4-8細胞期)到早期原腸胚后不等。年輕的單個卵塊內發育階段變異很小。孵化和室外水槽均在6-8天后開始。在所有情況下,姐妹卵塊半(即有藻類和無藻類的半)的孵化首次出現在相同的采樣間隔。
對保存的卵塊(所有均在觀察到孵化時立即保存)中心切片的檢查顯示,跨卵塊和處理,來自更中心位置的胚胎處于更早的發育階段。在所有卵塊中都發現了胚胎發育的梯度,因此最外層的胚胎處于孵化階段,而最早期的胚胎最中心。在有藻類的卵塊中發現的最早發育階段是早期無殼面盤幼蟲;該階段在實驗室卵塊中占30%,在室外水槽卵塊中占14%。對于無藻類的處理,最早階段是擔輪幼蟲,擔輪幼蟲存在于60%的實驗室卵塊和30%的室外水槽卵塊中。對于兩個實驗,與無藻類的對應半相比,在有藻類水中培養的卵塊半在孵化開始時具有更高比例的有殼胚胎(實驗室:t(9)=9.28,P<0.0001;室外:t(6)=8.83,P<0.0001)。平均而言,對于有藻類的卵塊橫切面,98%的胚胎有殼,而無藻類處理中的胚胎比例為59-66%。

圖1正在孵化的Siphonaria卵塊的橫截面,顯示從卵塊邊緣附近準備孵化的胚胎(左)到朝向卵塊中心的早期無殼面盤幼蟲(右)的發育梯度。箭頭指示有殼面盤幼蟲(SV)和無殼面盤幼蟲(PV)的示例。來自實驗室實驗的對照卵塊,無藻類培養。比例尺200μm
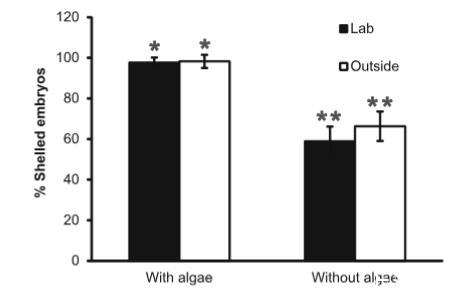
圖2孵化開始時為有殼面盤幼蟲的胚胎平均百分比(±95%置信區間),來自兩種處理和兩種不同的實驗條件(實驗室受控條件或室外變化條件)。星號表示每種條件下處理間的顯著差異(t檢驗,兩種情況下P<0.0001)。實驗室:n=10,室外:n=7
藻類對卵塊氧飽和度的影響
在第2天和第6天,藻類存在與光照處理之間存在顯著的交互作用(第2天:F1,23=37.70,P<0.0001;第6天:F1,25=49.3,P=0.01)。在光照下,有藻類的卵塊半具有更高的中心氧濃度,而無藻類的配對半則較低,但在黑暗下,所有有藻類或無藻類的卵塊中心氧濃度均低。無論光照如何,無藻類培養皿中水的平均氧飽和度相近且接近常氧(黑暗:98.0%,SD=1.1%;光照:97.5%,SD=1.4%)。黑暗中有藻類培養皿中的水為低氧(平均:46.0%飽和度,SD=12.3%),而在光照下為高氧(平均:264.1%飽和度,SD=52.3%)。平均而言,對于有藻類且在光照下的培養皿中的卵塊,年輕卵塊(第2天)的中心氧飽和度比老卵塊(第6天)高3倍(t(8)=3.86,P=0.005)。
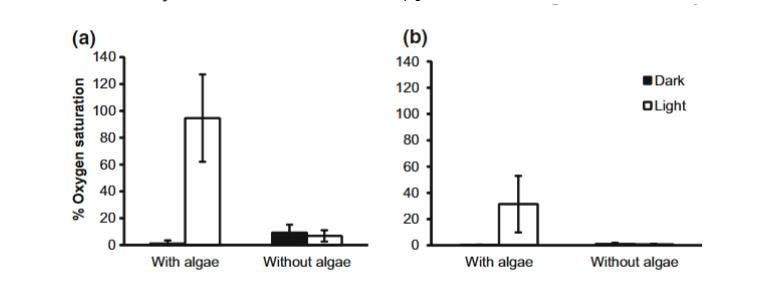
圖3兩種不同藻類處理以及經過8小時黑暗或4小時光照后卵塊中心氧飽和度的平均值(±95%置信區間),a為2天齡時,b為6天齡時。這些卵塊來自實驗室受控條件
潮池中的環境溶解氧在白天為飽和度的179%,夜間為飽和度的46%。來自該池的早期卵塊(前面盤幼蟲和早期面盤幼蟲階段)在白天平均中心氧飽和度為141.7%(SD=29.7%,n=10),夜間為2.2%(SD=2.3%,n=9)。
 相關新聞
相關新聞