熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
潮池氧飽和度與卵塊氧飽和度的關系
從白天測量的野外池中(溫度:13.9-17.5°C,n=6),前面盤幼蟲階段的卵塊中心氧飽和度顯著高于早期和晚期面盤幼蟲階段的卵塊(F2,28=8.93,P=0.001,分別為166.6%對比115.0%和116.1%;Tukey檢驗:P<0.05),并隨池氧飽和度增加而增加(F1,28=100.36,P<0.0001)。池氧飽和度解釋了前面盤幼蟲卵塊中心氧飽和度變異的68%,早期和晚期面盤幼蟲卵塊的90%和87%,且變化范圍從112.4%(SD=3.4%)到245.5%(SD=3.0%)。
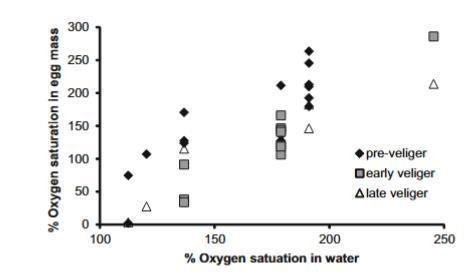
圖4潮池水中的氧飽和度驅動池中卵塊中心的氧飽和度(ANCOVA,P<0.0001),適用于包含多數胚胎處于三個不同發育階段之一的卵塊(ANCOVA,P<0.001):前面盤幼蟲黑色菱形(n=15),早期面盤幼蟲灰色方塊(n=11),以及晚期面盤幼蟲空心三角形(n=6)
我們在低潮時在一個晴朗的夏日測量了另外11個與海洋交換隔離的小潮池的氧飽和度,我們定性觀察到這些池的大型藻類覆蓋度不同。氧飽和度從無可見大型藻類的池的118%(SD=2.1%)到有豐富藻類(主要是石莼屬)的池的243%(SD=29.6%)不等。
討論
長期以來,人們認識到水生環境中的低氧可能對一系列進行底棲發育的類群產生毀滅性影響。但關注較少的是大型植物光合作用提供的氧氣在減輕這種風險方面可能起到的關鍵作用。在本研究中,與大型植物關聯增加了光照條件下底棲卵塊發育期間的氧飽和度。雖然藻類的存在不影響卵塊最外層胚胎的發育長度,但與藻類一起培養的卵塊在孵化開始時具有更高比例的晚期幼蟲。在所有情況下,無論藻類是否存在,配對卵塊半的孵化同時出現,這可能是因為從卵塊周圍水到最外層胚胎的短擴散距離使得這些胚胎能夠獲得足夠的氧氣,而與藻類存在無關。如果藻類暗呼吸消耗氧氣減緩了胚胎發育,這些影響是微妙的,表明該物種的胚胎能夠將其耗氧量調節到相當低的水平。相比之下,Fernandes&Podolsky(2011)報道了與鰻草關聯時腹足類Haminoea vesicula的孵化加速。由于該研究未直接測量氧氣,無法評估這種不同的反應是否表明低氧發育影響的物種特異性差異或其鰻草處理中光合作用和呼吸作用的對比動態。此外,在該研究中,孵化時間評估為50%胚胎孵化時的天數,而在本研究中,我們在孵化開始時停止了實驗。
Siphonaria australis卵塊在孵化開始后不久開始分解;失去結構完整性后,水流將分解卵塊并釋放剩余的胚胎。一旦從卵塊中釋放,尚未準備好孵化或開始浮游生活的胚胎——特別是那些尚未形成殼或游泳結構的胚胎——可能面臨更高的死亡風險。我們在所有卵塊中都看到了發育梯度,但當卵塊在無藻類條件下培養時,這種模式顯著更強;值得注意的是,對照處理(無藻類)中50%的卵塊在其最外層的兄弟姐妹孵化時,其中心胚胎處于最早的胚胎階段(即擔輪幼蟲)。相比之下,與藻類一起培養的對應卵塊中,幾乎所有中心胚胎都已達到更高級的有殼階段。這些梯度并非實驗室人為現象,因為在從野外收集的所有S.australis卵塊中也觀察到了發育梯度,并且已知存在于其他物種中。藻類光合作用提供的氧氣顯著增加了中心胚胎可用的氧氣量,并且可能是減少有藻類存在下卵塊發育梯度陡峭度的主要因素。在自然界中,這將允許更高比例的胚胎在卵塊分解前達到有殼面盤幼蟲階段。由于較大的卵塊往往更受氧氣限制(其他條件相同),藻類增加氧氣的好處將受卵塊大小的調節,藻類對大型卵塊更有益。
在實驗室中,老卵塊(第6天)的中心氧水平低于年輕卵塊(第2天),無論處理如何。類似地,在野外,盡管水中的氧飽和度直接驅動卵塊的中心氧濃度,但晚期卵塊的整體飽和度低于早期卵塊,這可能是因為隨著胚胎年齡增長代謝活動增加。不僅老卵塊通常更缺氧,而且許多類型的軟體動物胚胎隨著發育進展往往對低氧更敏感,可能是由于表面積/體積比減小和/或隨著胚胎發育代謝需求增加。因此,大型藻類的存在可能對老卵塊比年輕卵塊更有益。類似地,大型藻類可能對在卵塊或膠囊中完成變態的直接發育物種的卵塊產生更實質性的影響。
無論年齡或大小,無藻類保存的卵塊(對照)的中心總是缺氧的,并且在胚胎發育超過擔輪幼蟲階段后完全缺氧。當卵塊與藻類一起保存時,中心氧隨光照條件變化。當周圍水因數小時藻類光合作用而高氧時,卵塊中心的氧飽和度范圍從新產卵塊的95%飽和度到老卵塊的30%飽和度。經過模擬夜晚(黑暗中數小時)后,與藻類一起保存的卵塊的中心O2濃度與對照相似。這些結果與野外數據密切對應,其中一個潮池及其內部卵塊的氧飽和度也范圍從白天的高氧到夜間的低氧。然而,黑暗中藻類誘導的低氧對發育速率的任何影響似乎完全被白天光合作用獲得氧氣的好處所抵消。
野外潮池中的氧飽和度高度可變,既有時間上(由于晝夜光周期、溫度變化、生物代謝活動以及白天光照程度),也有空間上池與池之間,甚至池內。我們研究中的所有池都相對較小且淺,池與池之間的變異可能主要由生物因素驅動,特別是大型植物的相對豐度以及這些和其他生物(微生物、無脊椎動物、魚類)的代謝耗氧。作為池間變異的一個例子,在一個溫暖晴朗的夏日,我們發現一些池接近常氧,而其他池的氧水平約為飽和度的2.5倍。卵塊在高氧和低氧池中均有發現,這表明對于S.australis,潮間帶環境是一個復雜的景觀,后代可能根據父母選擇產卵的位置而從藻類中受益或不受益。雖然一些腹足類物種對卵塊沉積地點有選擇性,但據我們所知,尚無底棲海洋無脊椎動物的研究關注環境氧氣可用性作為父母選擇產卵地點的標準。澳大利亞和新西蘭的Siphonaria物種在高溫、干燥和紫外線輻射條件下并不避免產卵。
我們的氧氣測量表明,在自然界中,并且在整個發育過程中,Siphonaria胚胎常規經歷環境氧氣可用性的廣泛波動,從缺氧到2-3倍飽和度。對于單個胚胎,氧氣供應將取決于卵塊沉積地點和其在卵塊內的位置。對于靠近卵塊外圍的胚胎,氧水平在整個發育過程中可能接近常氧,或者可能在中等低氧和高氧之間交替。相比之下,中心胚胎可能無論沉積地點如何都受到氧氣限制,并且在無大型藻類時可能經歷長時間的完全缺氧。盡管我們未測試高氧和低氧對幼蟲性能的亞致死影響,但大多數成熟的S.australis胚胎在孵化時似乎功能完全,表明它們對兩者都有高耐受性。潮間帶海洋軟體動物通常表現出在面臨低氧或缺氧時代謝下調或停滯的高能力,而淡水肺螺類胚胎可以承受長時間的近缺氧條件。然而,尚未在軟體動物胚胎中研究成人代謝下調機制的存在。類似地,雖然軟體動物具有許多對抗高氧的生理防御機制,并且高氧已被證明至少對一種軟體動物的幼蟲生長產生負面影響,但總體而言,這些機制在胚胎中受到的關注較少。S.australis胚胎在其自然棲息地經歷劇烈的時空氧氣供應變化,因此這些和其他在潮池中發育的生物將是理解水生生態系統中低氧和高氧耐受代謝適應的有趣焦點物種。
有大量關于水生動物對低氧的生理和行為反應的文獻。然而,迄今為止,對可變氧氣條件的后果或驅動因素關注較少。大型植物是水生棲息地的基本組成部分,在決定氧氣變異性方面起著關鍵作用,特別是在更小、更孤立或短暫的棲息地中。我們已經證明,水生大型藻類提供的氧氣可以對底棲胚胎產生積極影響。大量類群產卵塊;氧氣對成功發育至關重要,并且通常對大多數內部胚胎有限制,這些胚胎應對非生物脅迫的能力很小。此外,低氧條件在許多水生棲息地中常見,并且隨著全球變化在某些地方日益加劇。因此,大型植物與水生動物之間以氧氣為媒介的關系可能對一系列類群而言是重要但尚未得到充分認識的有益相互作用。
致謝
本研究得到新西蘭皇家科學院國際科學與技術聯系基金對作者們的資助以及美國國家科學基金會的支持。
 相關新聞
相關新聞