熱線:021-66110810,66110819
手機(jī):13564362870
熱線:021-66110810,66110819
手機(jī):13564362870
結(jié)果
由于我們之前在分離純化的視桿細(xì)胞外段盤膜中發(fā)現(xiàn)了活躍的ATP合酶,并且其他人報(bào)道多酚類植物化學(xué)物質(zhì)作為線粒體ATP合酶和ATP酶活性的抑制劑,我們 prompted 測試其中一些化合物對視桿細(xì)胞外段中ATP產(chǎn)生或水解的影響。牛視網(wǎng)膜視桿細(xì)胞外段以完整視桿細(xì)胞外段的形式純化,仍包含胞質(zhì)、質(zhì)膜和盤膜囊泡,如同體內(nèi)狀態(tài)。我們之前對視桿細(xì)胞外段組分的表征排除了線粒體和內(nèi)段的污染,并且最近的數(shù)據(jù)證明純化的視桿細(xì)胞外段通過線粒體外的氧化磷酸化消耗氧氣并合成ATP。
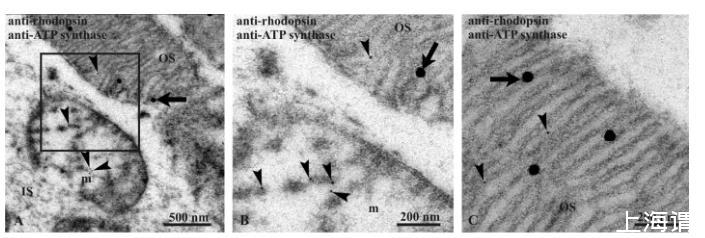
圖1、牛視網(wǎng)膜透射電鏡圖,檢測ATP合酶表達(dá)。(A-B) 用抗視紫紅質(zhì)抗體(大,40納米直徑金顆粒;箭頭)和抗ATP合酶β亞基抗體(小,10納米直徑金顆粒;箭頭)雙標(biāo)記的牛視網(wǎng)膜。(A)中的方框區(qū)域在(B)中放大,顯示視桿細(xì)胞外段(OS)中視紫紅質(zhì)和ATP合酶信號共定位的細(xì)節(jié),以及相鄰內(nèi)段(IS)中的線粒體(m),該處僅表達(dá)ATP合酶。(C) 一個(gè)視桿細(xì)胞外段的細(xì)節(jié)。
在此,我們進(jìn)行了牛視網(wǎng)膜的免疫金TEM成像。在視桿細(xì)胞外段中發(fā)現(xiàn)了視紫紅質(zhì)與ATP合酶β亞基的共定位(圖1A)。圖1B是雙標(biāo)記視桿細(xì)胞外段部分的放大圖,顯示視紫紅質(zhì)信號(40nm直徑金顆粒)與ATP合酶信號(10nm直徑金顆粒)的共定位。圖1C顯示同一雙標(biāo)記視網(wǎng)膜內(nèi)段的線粒體,在圖1D中放大,顯示僅存在ATP合酶信號。
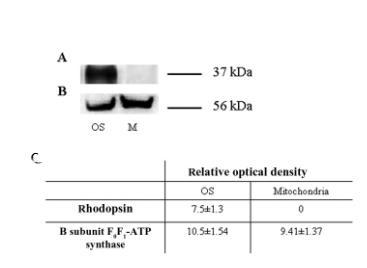
圖2、通過半定量Western印跡分析表征分離的視桿細(xì)胞外段。(A和B) 使用抗視紫紅質(zhì)(Rh)和抗ATP合酶β亞基抗體進(jìn)行的半定量Western印跡。樣品來自分離的視桿細(xì)胞外段(OS)和線粒體富集部分(M)。(C) 使用ChemiDoc軟件(Bio-Rad Laboratories)進(jìn)行的光密度分析。每個(gè)面板代表至少五次實(shí)驗(yàn)。
圖2中的Western印跡分析數(shù)據(jù)證實(shí)了圖1所示的結(jié)果,即視紫紅質(zhì)存在于視桿細(xì)胞外段中(圖2A),但不存在于用作陰性對照的線粒體中。ATP合酶β亞單位同時(shí)存在于視桿細(xì)胞外段和線粒體中(圖2B),光密度分析證實(shí)了這一點(diǎn)(圖2C)。
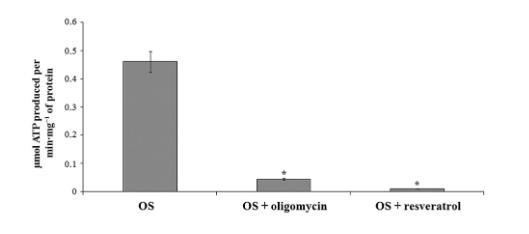
圖3、使用寡霉素和白藜蘆醇在純化的視桿細(xì)胞外段中進(jìn)行ATP合成。柱狀圖顯示在37°C,pH 7.3條件下,視桿細(xì)胞外段(40 μg·mL?1)在1分鐘內(nèi)的ATP生成量。加入10 μM寡霉素和30 μM白藜蘆醇分別抑制了90%和98%的ATP產(chǎn)量。數(shù)據(jù)顯示為平均值±標(biāo)準(zhǔn)差,n=4。*P < 0.01,配對Student t檢驗(yàn)。
ATP合酶在視桿細(xì)胞外段中具有催化活性,圖3顯示了視桿細(xì)胞外段勻漿的ATP合成。在存在NADH、琥珀酸鹽和ADP的情況下,檢測到最大活性為每分鐘每毫克蛋白質(zhì)產(chǎn)生0.430±0.060微摩爾ATP。ATP合成是特異性的,因?yàn)樗还衙顾兀?0%)抑制,寡霉素是經(jīng)典的線粒體ATP合酶抑制劑。這種ATP合成也被白藜蘆醇抑制(98%,圖3)。
為了測試白藜蘆醇損害盤膜質(zhì)子電位的能力,該化合物也用于先前用TMRM或JC-1(兩種線粒體膜電位探針)標(biāo)記的視桿細(xì)胞外段的細(xì)胞流式分析。表1報(bào)告了在存在NADH和琥珀酸鹽或加入白藜蘆醇后,使用視桿細(xì)胞外段勻漿獲得的結(jié)果。對照顯示TMRM和JC-1的均勻信號,表明存在膜電化學(xué)電位,如早期報(bào)道。與白藜蘆醇孵育30分鐘并未降低膜電位。在不同時(shí)間點(diǎn)進(jìn)行的測量長達(dá)1小時(shí)未顯示膜極化顯著降低。
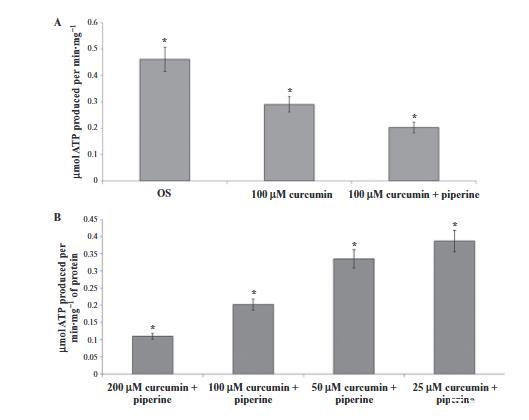
圖4、使用姜黃素和胡椒堿在純化的視桿細(xì)胞外段中進(jìn)行ATP合成。柱狀圖顯示在37°C,pH 7.3條件下,視桿細(xì)胞外段(40 μg·mL?1)在1分鐘內(nèi)的ATP生成量。(A) 加入100 μM姜黃素以及100 μM姜黃素加100 μM胡椒堿抑制了ATP產(chǎn)量。(B) 加入200、100、50或25 μM的姜黃素和胡椒堿組合抑制ATP合成。數(shù)據(jù)顯示為平均值±標(biāo)準(zhǔn)差,n=4。*P < 0.01,配對Student t檢驗(yàn)。
姜黃素單獨(dú)使用或與胡椒堿聯(lián)合使用對視桿細(xì)胞外段ATP產(chǎn)生的影響顯示在圖4A中。姜黃素(100μM)單獨(dú)使用時(shí)抑制ATP合成,但與胡椒堿(也使用100μM)聯(lián)合使用增加了抑制。這種組合(使用相等濃度)的效果是濃度依賴性的(圖4B)。
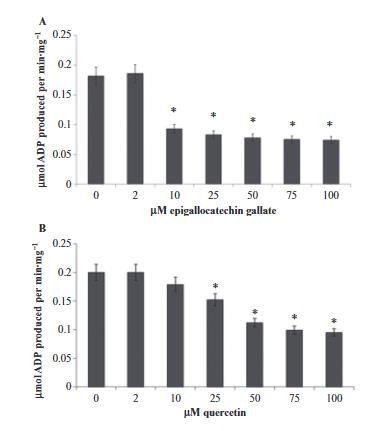
圖5、使用EGCG和槲皮素在純化的視桿細(xì)胞外段中進(jìn)行ATP水解。柱狀圖顯示在37°C,pH 7.3條件下,視桿細(xì)胞外段(40 μg·mL?1)在1分鐘內(nèi)的ATP水解量。(A) 在不同濃度下,EGCG或(B)槲皮素抑制視桿細(xì)胞外段制備物中的ATP水解。數(shù)據(jù)顯示為平均值±標(biāo)準(zhǔn)差,n=4。*P < 0.01,配對Student t檢驗(yàn)。
 相關(guān)新聞
相關(guān)新聞