熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
視覺皮層
外側膝狀體(LGN)的小放大倍數和簡單的功能組織使其成為量化氧反應空間特性的理想模型,但其他視覺區域是否表現出相似特性?盡管視覺皮層的放大倍數大于LGN且功能組織更為復雜,但通過利用該區域的柱狀組織結構,可以系統性地改變神經活動的空間模式。在先前研究中,我們證實組織氧濃度的變化可用于預測微電極附近神經元的方位選擇性和眼優勢。本文通過兩個示例和這些數據的群體統計(圖6),使其可與LGN的氧反應進行對比。
在圖6中,左側圖標代表用于視覺皮層記錄位點的個體優勢眼與非優勢眼刺激條件(圖6第一列),右側圖表則顯示相應的神經反應與氧反應(圖6第二至四列)。需注意,特定記錄位點實際采用的刺激方向組合取決于所記錄神經元的方位調諧特性(參見材料與方法部分的實驗方案)。圖6A展示了與記錄神經元偏好方向正交的光柵刺激所引發的反應。該刺激推測能在方位柱中引發強烈的神經反應一這些方位柱靠近但未重疊于我們傳感器的尖端。雖然神經活動模式并不完全一致,但這種正交刺激類似于LGN實驗中采用的小型掩膜刺激。兩種刺激均能引發靠近但未重疊于記錄位點的神經活動。
負氧反應明確地局限于神經活動發生的部位。
這一結果在我們所有的LGN記錄位點群體中也保持一致。圖5以與圖3相同的格式展示了群體數據。最大負向變化(水平軸)隨著刺激物遠離RF而遞減。組織氧含量的正向變化相對于大視野刺激期間觀察到的變化較小。隨著刺激物遠離RF,平均正向變化略有增加,但這可能歸因于微小的活動依賴性變化,或基線噪聲與缺乏強烈負向變化的綜合作用。雖然我們在圖2和圖3中展示了RF外部的刺激能引發組織氧含量的顯著正向變化,但那些大視野刺激引發的神經活動總量遠大于圖4和圖5中小型刺激引發的活動量。這很可能是大型刺激引發顯著正向氧變化的根本原因刺激點。氧氣對這些刺激的反應也相似。兩者都會引起氧氣的顯著正向變化,而初始負向變化很小或沒有。圖6B-F顯示了另外五種以給定記錄點(圖6D)最佳值為中心的方向條件下的氧氣和神經反應。這些刺激可能在接近和重疊傳感器尖端的方向柱內引起神經活動。刺激方向越接近最佳值,初始負向變化越大,延遲正向變化越小。這一結果與我們在LGN中的測量結果一致,其中與我們的傳感器重疊的神經活動通常引起組織氧氣的負向變化,而周圍區域的活動引起略微延遲的正向變化。最后兩個刺激條件顯示了向主導眼(圖6G)和非主導眼(圖6H)呈現的最佳方向大視野刺激的氧氣和神經反應。
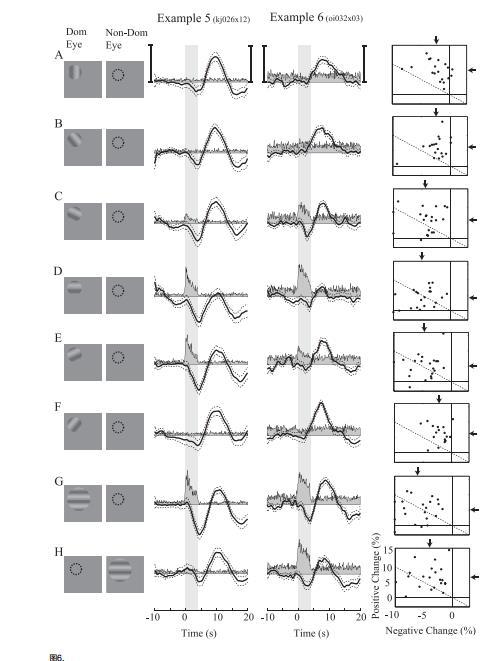
圖6.視覺皮層中氧反應與神經元方位選擇性和眼優勢的調諧特性。A-H,視覺皮層神經活動與氧測量的刺激響應配對。首列展示優勢眼與非優勢眼的漂移光柵刺激。虛線圓圈表示感受野(RF)的位置與尺寸。 A-F,方位調諧:呈現六種光柵方向。正交(A)與最優(D)方向的光柵與另外四個方向(B,C,E,F)交替呈現,這些方向被選用于粗略采樣各神經元的方位調諧曲線。 G-H,眼優勢:兩個大視野最優方向漂移光柵(雙眼各一)與方位條件交替呈現,以觀察由眼優勢引起的差異。第二、三列分別顯示 80次和 48次試驗平均的多單元神經活動(灰色填充)與氧響應(黑色)。虛線表示平均氧響應的 ±1 標準誤。陰影區域標示刺激起始與持續時間。垂直刻度標尺:示例5左為 12次/秒峰值放電率,右為 10%氧濃度變化;示例 6左為 17次/秒峰值放電率,右為 16%氧濃度變化。第四列顯示皮層記錄位點群體(n=21)的最大負向與正向氧變化。除垂直刻度外,圖示規范與圖3相同。
第一個示例點具有很強的單眼部刺激。優勢眼中的最佳刺激能引發強烈的神經響應、組織氧含量顯著負向變化以及延遲的正向氧變化(圖6G,第2列)。同一刺激作用于非優勢眼時,在我們傳感器附近僅引發微弱神經響應,但推測激活了數百微米外鄰近的眼優勢柱。該刺激的氧響應特征為小幅負向變化和相對較強的正向變化(圖6H,第2列),與非最佳方位角條件相似。第二個記錄點顯示出雙眼神經響應特性(圖6G、H,第3列)。
貓視覺皮層中大多數神經元具有雙眼特性,因此無論哪只眼睛接收最佳刺激...
刺激單側眼球會引發強烈的神經響應。在此示例中,氧合反應對左右眼刺激均呈現雙相響應特征(圖6G、H,第三列)。左右眼刺激條件下響應幅度的微小差異可能源于兩種刺激所引發的神經活動模式不同。但由于該傳感器相對于眼優勢柱布局的位置尚不明確,因此難以確定這種差異的具體成因。
圖6第四列展示了我們視覺皮層21個記錄位點的正負氧反應強度,采用與圖3和圖5相同的呈現格式。所有位點的氧反應模式均與圖6所示的兩個示例及外側膝狀體的測量結果一致。平均負向氧反應(垂直箭頭)在優勢眼接收最優朝向刺激時達到峰值,而平均正向氧反應(水平箭頭)則在非最優刺激及非優勢眼接收刺激時最為顯著。
將神經活動與組織氧變化關聯的模型
盡管我們組合的氧和神經測量是在組織中的單個點獲得的,但上述結果表明,在解釋組織氧的變化時必須考慮傳感器視野之外的神經響應。為此,我們開發了一個簡單的線性模型。該模型將神經活動的時空模式與我們的傳感器測量的組織氧的活動依賴性變化聯系起來。對于該模型,我們假設組織氧的變化由兩個時間響應分量的線性總和決定:
R(ro, t) = P(ro, t) + N(ro, t) (1)
其中t是時間,ro是我們微電極傳感器在腦組織中的位置,R是氧響應的時間過程,P和N分別是正向和負向響應分量。注意,R表示與給定神經響應相關的氧變化,不包括與基線狀態相關的持續變化。氧響應的這種雙分量模型的動機是本文和先前報告中描述的實驗測量結果。P和N通過將時空神經響應A(r, t)與獨立的時空脈沖響應函數Sp(r, t)和Sn(r, t)進行卷積來確定:

其中hp和hn是時間脈沖響應函數,wp和wn是具有獨立增益(gp, gn)和寬度(σp, σn)參數的高斯PSF。σ決定了神經活動被積分的體積,g對PSF的幅度進行縮放。參數σp和σn尤為重要,因為它們決定了神經和氧響應之間的空間特異性。
